
第三篇 基因信息的传递
第十一章 DNA的生物合成(复制)
第一节 半保留复制
一.相关概念
(一)复制(replication)(DNA的生物合成)
脱氧核糖核酸的复制(replication of deoxyribonucleic acid)。遗传信息从亲代DNA传递到子代DNA分子上的的过程。即以亲代DNA分子为模板合成两个结构完全相同的子代DNA分子的过程。此过程同时将亲代DNA分子所储存的遗传信息准确地复制到子代DNA分子中。
DNA的忠实复制保证了生物性状的代代相传。
1.体外以DNA为模板、四种dNTP为底物,用DNA聚合酶可合成新生的DNA。
2.在细胞内DNA复制是一个由多种酶协同作用的复杂过程,包括:
(1)DNA复制的起动;(2)DNA双螺旋的解开;(3)新生DNA链的合成;(4)复制后DNA的修饰;(5)受损伤的DNA的修复;(6)复制的调节控制等方面。
(二)转录transcription(RNA的生物合成)
在生物细胞内以DNA为模板合成与DNA某段碱基序列互补的RNA分子的过程。
此过程将遗传信息传递到RNA分子中(主要是mRNA分子)。以DNA为模板合成RNA的过程就是RNA的生物合成。
此当然包括各种RNA分子,但只有mRNA向下传递遗传信息。
(三)翻译translation)蛋白质的生物合成)
以mRNA为模板,将其中的碱基顺序(密码)转变为蛋白质中氨基酸的排列顺序的过程。即:在遗传密码的指导下合成蛋白质多肽链的过程。
(四)中心法则The central dogma
遗传信息在细胞内的生物大分子间转移的基本法则。
1.遗传信息:包含在DNA或RNA分子中的具有功能意义的核苷酸顺序。
2.遗传信息的转移:包括核酸分子间的转移、核酸和蛋白质分子间的转移。
1957年F.H.C.克里克最初提出的中心法则是:DNA→RNA→蛋白质
它说明:遗传信息在不同的大分子之间的转移都是单向不可逆的,只能从DNA到RNA(转录),从RNA到蛋白质(翻译)。这两种形式的信息转移在所有生物的细胞中都得到了证实。
1970年H.M.特明和D.巴尔的摩在一些RNA致癌病毒中发现它们在宿主细胞中的复制过程是先以病毒的RNA分子为模板合成一个DNA分子,再以DNA分子为模板合成新的病毒RNA。前一个步骤被称为反向转录,是上述中心法则提出后的新的发现。
1970年克里克重申了中心法则的重要性,提出了更为完整的图解形式(见图中心法则)。
3.遗传信息的转移分为两类:
(1)实线箭头表示:
这三种遗传信息的转移方向普遍地存在于所有生物细胞中。包括DNA的复制、RNA的转录和蛋白质的翻译,即①DNA→DNA(复制);②DNA→RNA(转录);③RNA→蛋白质(翻译)。
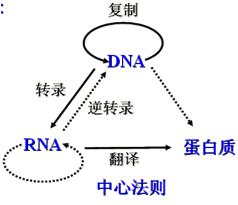
(2)虚线箭头表示:
是特殊情况下的遗传信息转移,包括RNA的复制,RNA反向转录为DNA和从DNA直接翻译为蛋白质。即①RNA→RNA(复制);RNA复制只在RNA病毒中存在。
②RNA→DNA(反向转录);最初在RNA致癌病毒中发现,后来在人的白细胞和胎盘滋养层中也发现了反向转录酶的活性。③DNA→蛋白质。仅在理论上具可能性,在活细胞中尚未发现。
4.克里克认为:图解中没有箭头指向的信息转移是不可能的,即
(1)蛋白质→蛋白质;(2)蛋白质→RNA;(3)蛋白质→DNA。
中心法则的中心论点是遗传信息一旦转移到蛋白质分子之后,既不能从蛋白质分子转移到蛋白质分子,也不能从蛋白质分子逆转到核酸分子。
克里克认为这是因为核酸和蛋白质的分子结构完全不同,在核酸分子之间的信息转移通过沃森―克里克式的碱基配对而实现。但从核酸到蛋白质的信息转移则在现存生物细胞中都需要通过一个极为复杂的翻译机构,这个机构是不能进行反向翻译的。
因此如果需要使遗传信息从蛋白质向核酸转移,那么细胞中应有另一套反向翻译机构,而这套机构在现存的细胞中是不存在的。
5.中心法则合理地说明了在细胞生命活动中两类大分子的联系和分工:
(1)核酸的功能――储存和转移遗传信息,指导和控制蛋白质的合成;
(2)蛋白质――主要功能是进行新陈代谢活动和作为细胞结构的组成成分。
二.复制的方式――半保留复制semiconservative replication
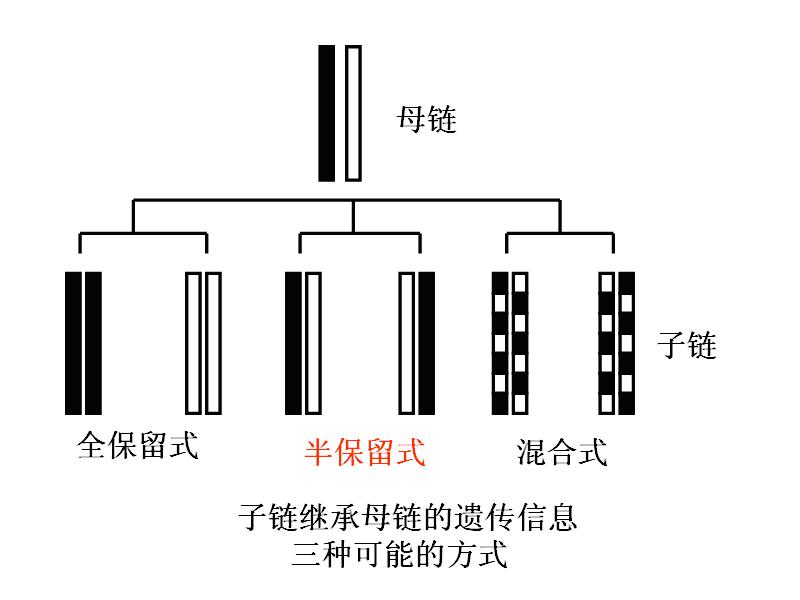
1953年J.D.沃森和F.H.C.克里克在阐明了DNA分子双螺旋结构之后,又提出了DNA半保留复制的假设:
(一)内容
1.亲代DNA的两条链可分别作为模板,各合成一条新生的DNA链。
2.子代DNA中一条链来自亲代,另一条链是新合成的。
3.基于碱基互补,两个子代DNA分子同亲代的碱基顺序完全一样。
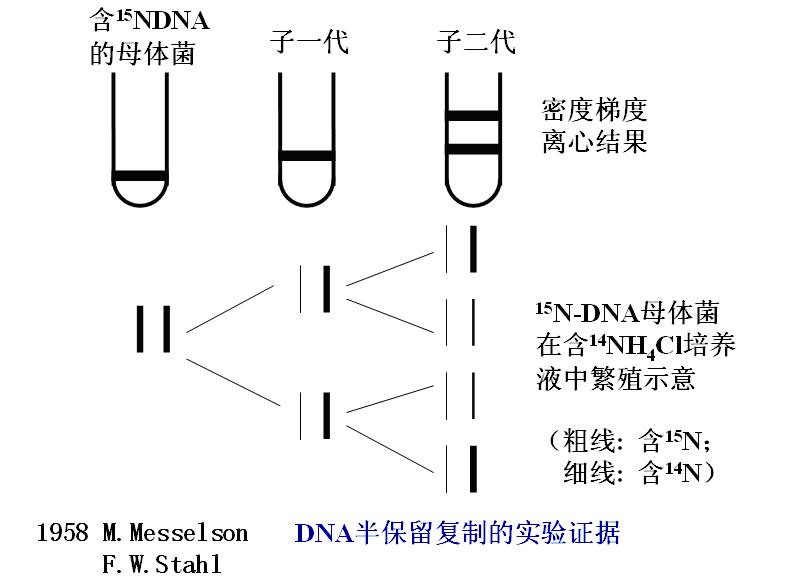
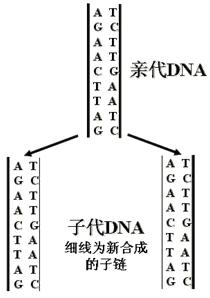
这个假设在1957年前后已为若干实验所证实(图: DNA的半保留复制)。
(二)实验原理和过程
1.原理
(1)细菌可以利用NH4Cl作氮源合成DNA。
把细菌放在15NH4Cl的培养液中培养若干代――分离出的DNA是含有15N的“重”DNA。密度比一般的含14N的DNA高。
(2)14N和15N的分布可用新发展的密度梯度沉降平衡技术来检测。
将少量DNA溶于密度接近于DNA浓度(约1.79g/cm3)的氯化铯浓溶液中。将此溶液离心直到接近平衡。
离心时沉降与扩散这两个相对抗的过程使氯化铯在离心池中形成了浓度梯度,结果是建立了一个范围为1.66―1.76g/cm3)的稳定的密度梯度。
此密度梯度中,DNA在离心力作用下停留在密度与DNA浮力密度相同的溶液位置中。
大分子量DNA表现出明晰的区带,可以通过紫外收收检测出来。由于14N-DNA和15N-DNA分子在密度上差异1%,它们可以经过密度梯度准确地分开。
2.过程
(1)把含有15N-DNA的细菌放回普通NH4Cl培养液中培养。
定期提取DNA作密度梯度分析(营养充足时20分钟可以生长成新一代)。
(2)实验结果证明(如图),15N-DNA是按几何级数被逐渐“稀释”的。
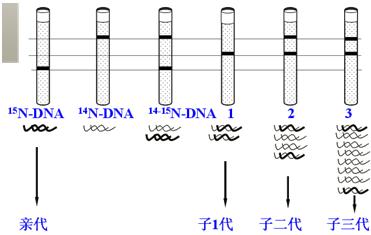
(三)半保留复制的意义
子代保留了亲代DNA的全部遗传信息,体现在代与代之间DNA碱基序列的一致性上。
第二节 DNA复制的酶学
一、复制的化学反应
核苷酸之间是通过磷酸二酯键的生成而逐一聚合的。反应的四种原料(底物)是dNTP而非dNMP(N=A、G、C、T的任何一种)。
每一步的生化反应:
(dNMP)n + dNTP ――→ (dNMP)n+1 + PPi
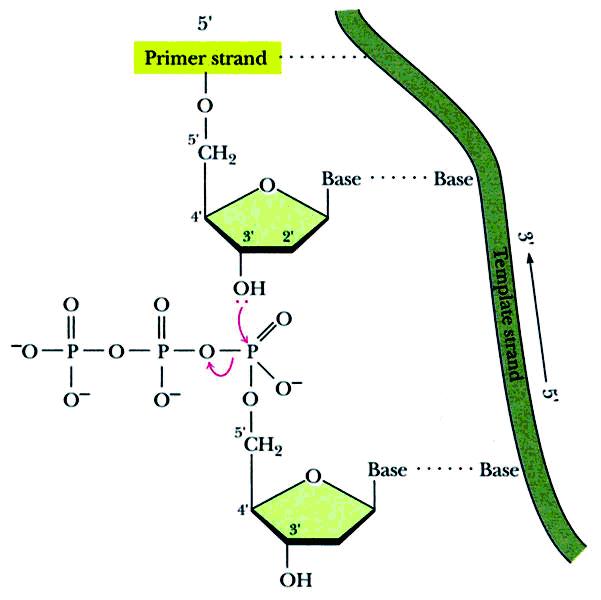
链的延长有方向性:
新链只能从5'-端向3'-端延长。底物的5'-P是加合到原有链的3'-OH上形成磷酸二酯键的。两条链的走向相反,但各自的合成方向相同,都是5'-端向3'-端延长。
见下图:复制过程中脱氧核苷酸的聚合。
二、复制中解链与DNA的拓扑学变化
(一)母链DNA模板(template)
1.两条链解开成单链后都可以作为模板。DNA只有解开成单链才能作模板。
2.DNA双螺旋的解开是一些酶和蛋白质协同作用的结果。
(二)解开超螺旋、解开双螺旋双链成单链的酶
目前已知的解旋、解链酶类,至少有三类。
其共同的作用是解开、理顺DNA链,维持DNA在一段时间内处于单链状态。
1.解螺旋酶(helicase)(解链酶、复制蛋白、rep蛋白、DnaB):使DNA的两条互补链分离。在复制叉前解开一小段DNA双链。在整个复制过程中此酶随复制叉的伸展而移动。此过程消耗能量,2ATP/bp。
大肠杆菌复制前DnaA辨认起始点、DnaC协助DnaB结合在起始点并打开双链。
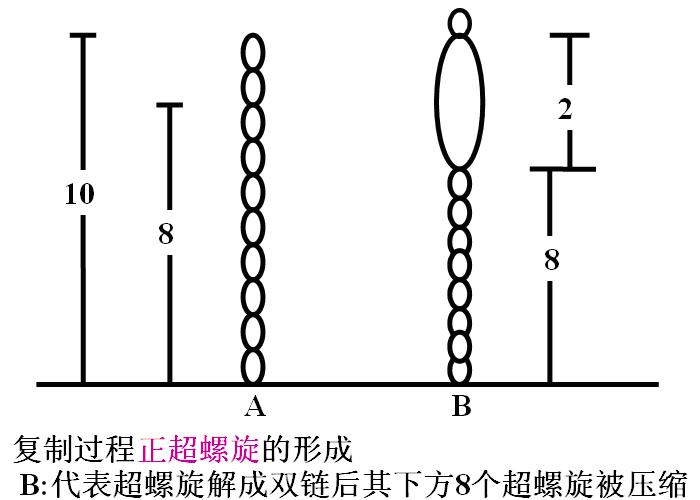
2.DNA拓扑异构酶(DNA topoisomerase)(拓扑酶):松弛DNA超螺旋,克服紧密扭结现象。(拓扑:物体或图象作弹性移位或变形而物体不变)。
复制过程超螺旋的形成见下图。

(1)拓扑异构酶I:切开DNA的单股,松弛正、负螺旋后在封闭缺口。不需要ATP参与。
①在大肠杆菌拓扑异构酶I最初发现,称为ω蛋白。
②在真核生物拓扑异构酶I曾用过多种名字:转轴酶(swivelase)、解缠酶(untwisting enzyme)、切口-封闭酶(nicking-closing enzyme)、松弛酶(relaxing enzyme)。
(2)拓扑异构酶II:同时切断DNA的双股,松弛螺旋后封闭变为负超螺旋。仅连接需要ATP参与。
拓扑酶对DNA分子的作用是可以水解、又可以连接磷酸二酯键。参与复制的全过程。
①在原核生物拓扑异构酶II又称为旋转酶(gyrase);
②在真核生物拓扑异构酶II分为几种亚型。
3.DNA单链结合蛋白(single stranded DNA binding protein)(DBP或SSB):
几个分子的DBP结合到DNA的单链上,防止DNA重新形成双螺旋而保持单链状态,同时防止单链模板被核酸酶水解。曾用名――螺旋反稳定蛋白(HDP)。
作用过程:结合――脱离――再结合,和聚合酶的沿模板向前移动不同。
三、引物酶和引发体
(一)引物酶与引物
1.引物酶primase :催化引物合成的酶,属于RNA聚合酶又不同于转录酶。与解螺旋酶(DnaB)共同作用。
2.引物:DNA新生链的合成都是以引物酶先合成一小段RNA链为起始的,这一段RNA称为引物。引物通常长10~50核苷酸。RNA引物最终被切除并由DNA取代。
DNA聚合酶只能延长而不能开始新的DNA链合成,需要引物――短的RNA。
(二)引发体primosome:
DnaA、DnaB、DnaC及其它复制因子形成复合体,再结合引物酶形成较大的聚合体再结合到模板DNA上,构成引发体。
引发体的下游解开双链,再由引物酶催化引物的合成。
四、DNA聚合酶
(一)全称:
DNA依赖的DNA聚合酶(DNA-dependent DNA polymerase )简写DNA-pol或pol)。
1956年A. Kornberg从大肠杆菌中首次分离到DNA聚合酶,因此这个酶又称为科恩伯格酶。又叫复制酶(repli-case)。后来证明此酶就是DNA聚合酶Ⅰ。
(二)作用:
1.聚合:以DNA为模板,以4种dNTP为底物,在引物存在下催化新的DNA链的合成。
2.外切:部分具有核酸外切酶作用,起校读和损伤修复的作用。对大肠杆菌的DNA聚合酶研究得较为透彻。
(三)种类
1.原核生物(大肠杆菌)包含3种DNA聚合酶:
(1)DNA聚合酶Ⅰ,其特点是:
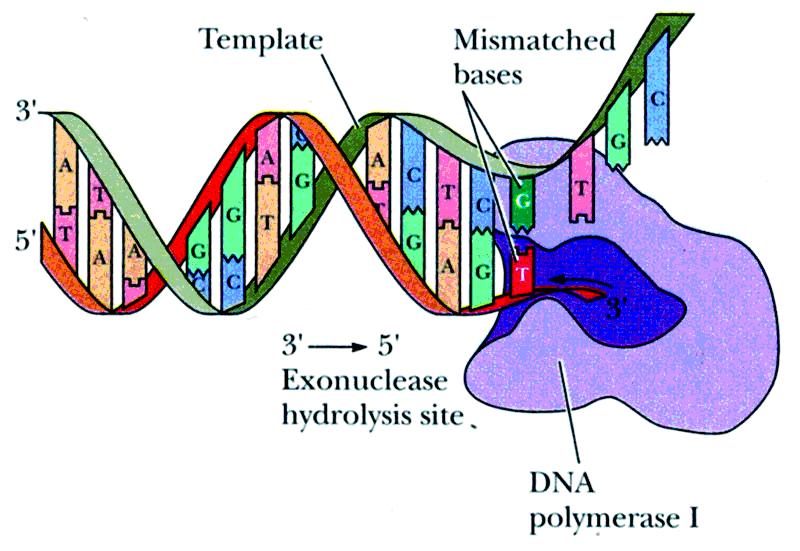
①DNA模板是必需的:不存在模板时,不表现聚合酶活力;
②引物也是必需的:DNA聚合酶只能从引物开始延长DNA链,而不能在DNA模板上重新合成一条新链;
③催化链的单向延长:以脱氧核苷三磷酸为底物,从5'→3'方向延长DNA链,而不催化3'→5'反应;每分钟催化20个核苷酸聚合。
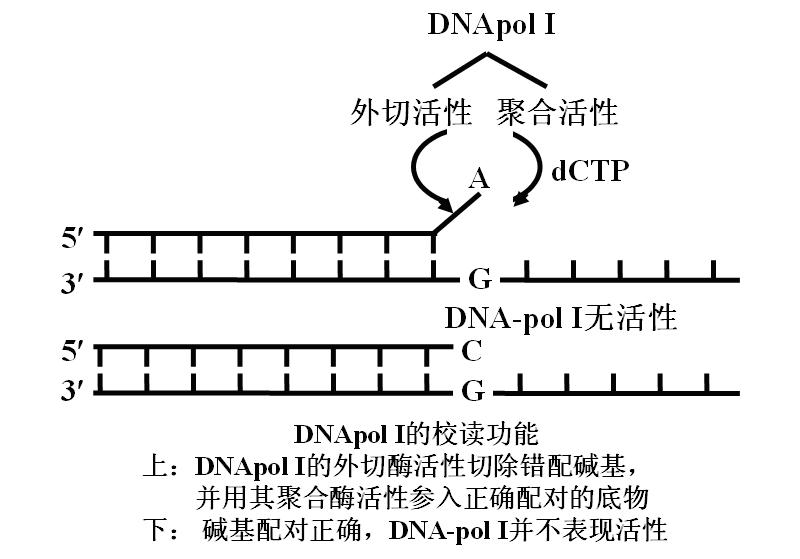
④具有3'→5'外切酶活力――大片段(Klenow fragment)的作用(常用做工具酶)
当新掺入的核苷酸同模板错配时,它能识别并切除不配对的核苷酸,然后才能进行下一步聚合反应,因此这个酶能够校正聚合过程中碱基的错配,保证DNA复制的高度精确性。
这是此酶在活细胞中的主要功能。它不是复制的主要酶。主要作用是“即时校读”。复制不出错误时不起作用。
⑤具有5'→3'外切酶活力――小片段的作用
DNA聚合酶Ⅰ的主要功能是切除引物,填补由此产生的缺口,并在DNA损伤修复中起作用,可以切除突变的片段。
(2)DNA聚合酶Ⅲ:
DNA聚合酶Ⅲ具有同DNA聚合酶Ⅰ相类似的特性。只有3'→5'一种方向的核酸外切酶活力。每分钟催化105个核苷酸聚合。
在细胞内,聚合酶Ⅲ同多种肽链组成复合物,称为DNA聚合酶Ⅲ全酶,可能它才是真正负责大肠杆菌染色体DNA复制的酶。
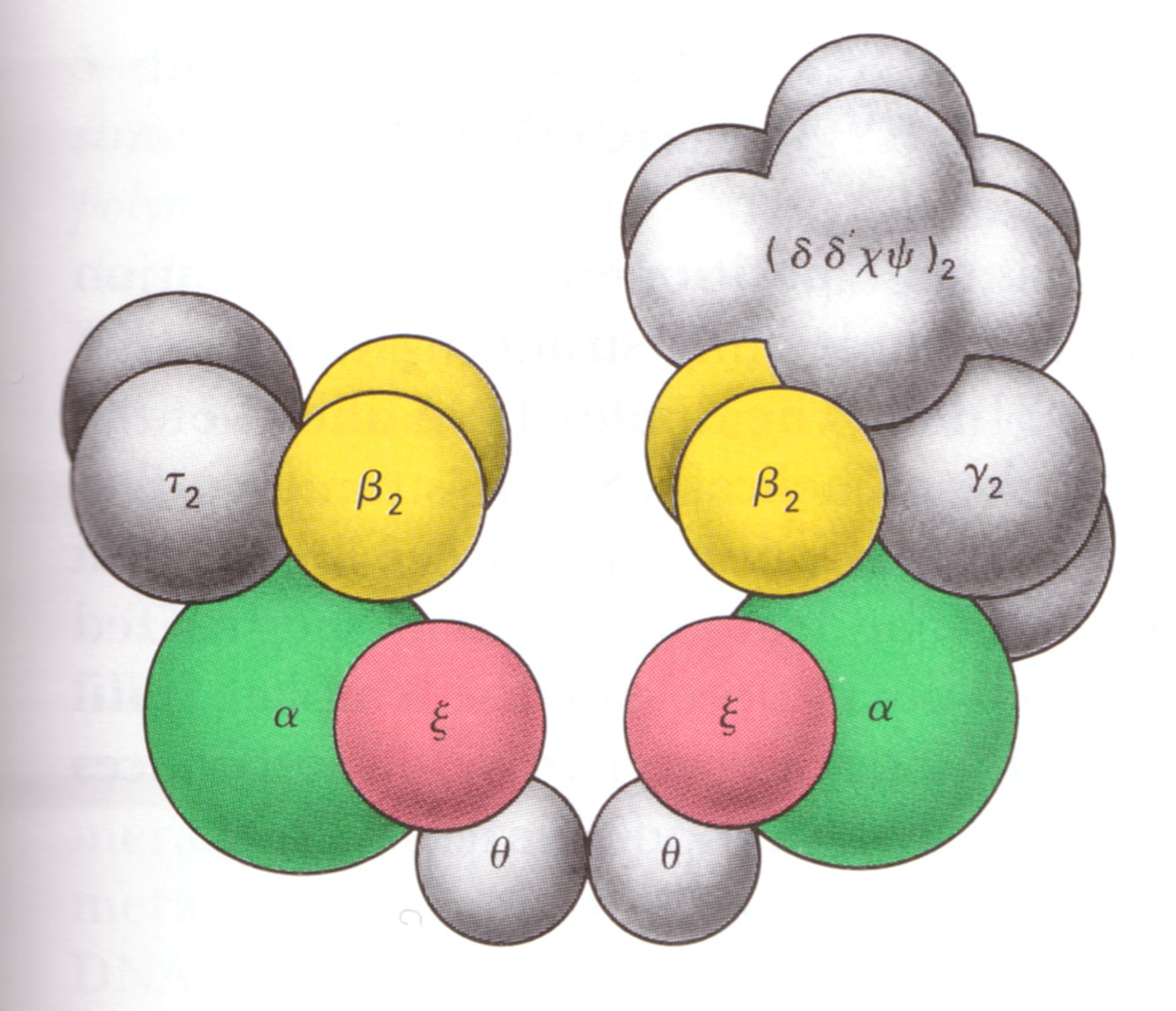
DNA聚合酶Ⅲ对核苷酸的掺入有选择性,可以保证复制的保真性。
(3)DNA聚合酶Ⅱ:
pol ⅡI只在其他两种酶缺如的情况下其作用。真正的功能不清楚。具有两种方向的核酸外切酶活性。
细胞内分子个数比例:DNApolⅠ:Ⅱ:Ⅲ = 400:40:20,但是pol III的比活性超过pol I的10倍。
2.真核细胞中通常含有5种DNA-pol,分别称为α、β、γ、δ、ε。
(1)DNA聚合酶α:存在于细胞核内,它的组成复杂而又不稳定,至今还没有完全纯化。鼠DNA聚合酶α至少由5条肽链组成。同大肠杆菌DNA聚合酶不同,真核生物DNA聚合酶α不具有外切酶活力。
该酶活力随着细胞周期而变化,在DNA合成时期,这个酶活力达到高峰,因此推测这个酶负责核DNA的复制。只能延长几百个核苷酸,故认为是延长随从链的。
(2)DNA聚合酶β:活力不随细胞周期而变化,它的功能主要在于修复受损伤的DNA。且只在没有其他DNA聚合酶时起作用。
(3)DNA聚合酶γ:可能就是早先发现的线粒体DNA(mitochondia DNA,mtDNA)聚合酶。
(4)DNA聚合酶δ:复制的主要酶。延长领头链。
(5)DNA聚合酶ε:与原核的DNApol I相似,在复制中起校读、修复、填补缺口的作用。

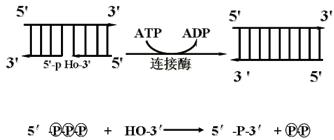
五、DNA连接酶(DNA-ligase)
复制过程中必需的酶之一。在模板存在下,它把DNA片段的3'-羟基末端同相邻片段的5'-磷酸通过磷酸二酯键连接起来。此过程需要ATP参与。
在岗崎片段的连接、DNA的修复和DNA重组中,DNA连接酶都是不可缺少的。
第三节 复制的过程
复制的过程包括起始、延长、终止三个连续阶段。(原核细胞为主,真核仅作对比)
一、复制的起始与起始点
(一)复制的起始点
复制只能在DNA分子上特定区域内起始,这一段特定的DNA顺序称为复制起始点。
1.原核生物染色体、病毒和核外DNA通常只有一个复制起始点。
(1)大肠杆菌复制起始点Oric(origin)在异亮氨酸、缬氨酸基因(ilv)附近。
它的必需的最小长度为245碱基对。具有专一顺序,富含A-T。
3组串联重复序列:1―13、17―29、32―44;两对反向重复序列:58―66←→166―174、201―209←→237―245。
复制开始此顺序首先被DnaA识别并且分开双链,另一个蛋白DnaB(解链酶,helicase)连接到双链上向两个方向解链。如图:
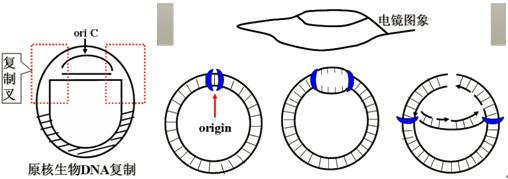
(2)双向复制(bidirectional replication):E.coli从固定的起始点开始同时向两个方向复制。
2.真核生物染色体具有很多复制起始点,同时形成许多复制单位,彼此相隔约3~10万碱基对。但是不一定同时启动。如图。
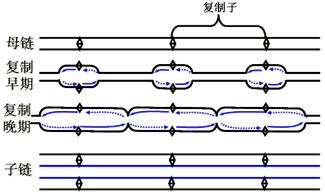
复制子(replicon):两个起始点之间的DNA片段,称为一个复制子。
(二)复制叉
按半保留复制的机制,DNA分子的两条链分别作为模板而进行复制,因此在复制起始点的两端就形成叉状结构,称为复制叉。(如图:大肠杆菌DNA复制叉及复制过程简图)。
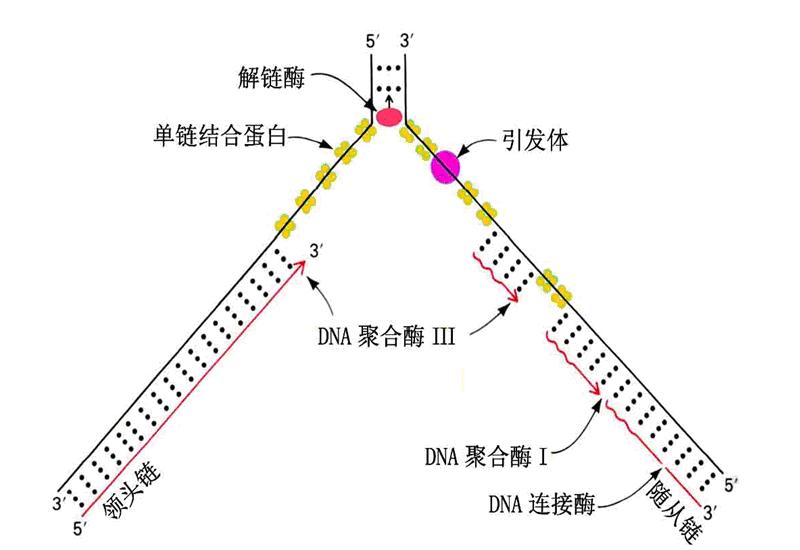
1.大多数生物随着复制的进行,两端的复制叉以反向等速方式前进。
2.某些染色体外的DNA复制叉前进的方式较为特殊(如噬菌体、质粒和线粒体等)
①双向而不等速前进,②仅单向前进,③滚筒方式前进,④两条新生链不同时复制。
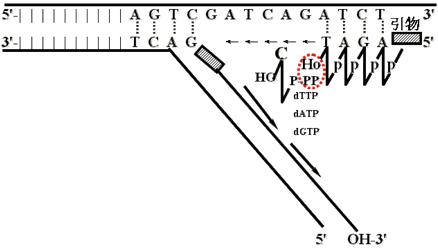
(三)岗崎片段
1.复制是有极性的
DNA聚合酶只能催化DNA链5'→3'方向延长(新链的生长方向),而不能催化反方向(即3'→5')的延长。被复制的链(模板)的走向是3'→5'。
2.复制是半不连续的
前导链(主导链、领头链)、随从链(滞后链)、冈崎片段
(1)同复制叉移动方向一致的一条新生链是连续合成的,称为前导链;
(2)同复制叉移动方向相反的一条链只能以非连续方式合成,称为滞后链。
岗崎首先发现这些短的非连续合成的新生片段,因此称为岗崎片段。
在引物被切除,缺口被补满以后,岗崎片段由DNA连接酶连接成一条长链。
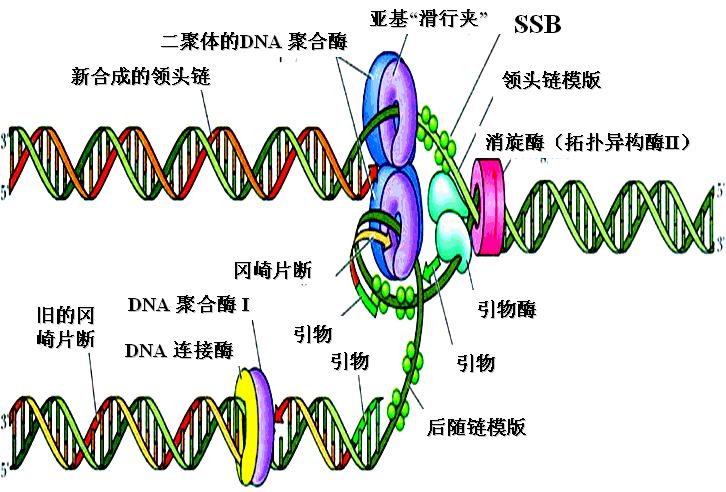
(四)复制起始过程(以大肠杆菌为例)
1.复制开始――引发体形成
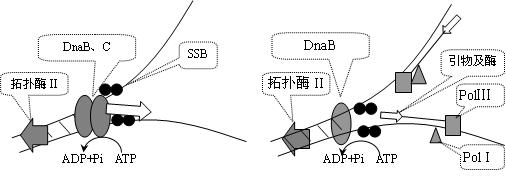
①解开螺旋(解链酶、拓扑异构酶、ATP)②形成复制叉(SSB)

引发体的蛋白部分在DNA链上移动、解链,同时逐步置换出DnaA蛋白,同时SSB参与单链的稳定。移动、解链的过程需要拓扑异构酶的理顺。
(3)合成引物5-100bp(引物酶或真核生物细胞polα)
移动到一定位置、双链解开足够复制的长度,新的游离核苷酸(NTP)以模板作依据掺入合成引物,带有3′-OH。
二、DNA片段形成――复制的延长
脱氧核糖单核苷酸(以dNTP为原料)逐个在模板的指引下(碱基配对)加入形成磷酸二酯键,使新链逐渐延长。
复制速度:大肠杆菌(3000kb)20分钟繁殖一代,相当于2.5kb/秒;真核可能较慢,但却是多个复制子同时进行。
1、前导链:聚合酶III(真核聚合酶δ)催化完成。
2、后随链:聚合酶III(真核聚合酶α)催化完成。
每隔约1000核苷酸引发一次新生链。然后RNA引物被DNA聚合酶Ⅰ切除,同时填满由此而产生的缺口,相邻的片段由DNA连接酶连接成为长的DNA链。
3、引物水解及其引物空隙填补
聚合酶I(真核核酸酶H)切除引物,同样聚合酶I(真核聚合酶β)填补引物水解所留下的空隙。
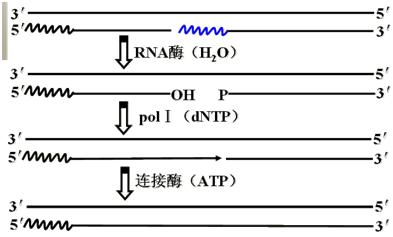
三、复制的终止――完整DNA单链的形成
连接酶催化连接两个DNA片段形成3'→5'磷酸二酯键。
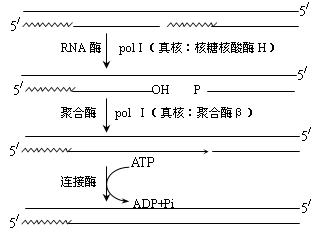
各个环节不是独立的,往往是同时进行的。所以各阶段并无严格划分。
四、复制的保真性
复制的保真性依赖pol的核酸外切酶活性及其严格的碱基配对。至少有三种机制:
1.遵守严格的碱基配对规律――模板。
2.聚合酶对碱基的选择功能――DNA-pol Ⅲ。
3.复制出错时的即时校读功能――DNA-polⅠ。
补充说明:
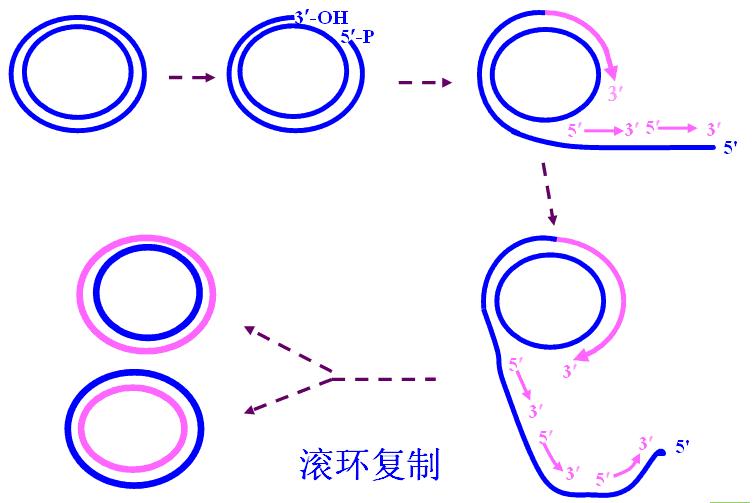
(一)滚环复制
简单低等生物的DNA、染色体外DNA采用滚环复制。
1.环状双链DNA先打开单链缺口、5/-端外伸
2.以外伸的单链作模板合成新链(不连续)。
3.没有开环的单股母链边滚动、边连续复制。
4.最后合成两个环状子双环。
滚环复制可能不需要引物。
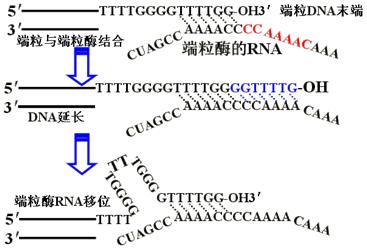
(二)真核生物的端粒和端粒酶
1.端粒:真核生物染色体线性DNA分子末端的膨大、粒状结构。像两顶帽子盖在染色体两端。端粒共同特点是富含G、C碱基的短序列并且多次重复。
仓鼠和人类的端粒DNA都有-TTAGGG-序列,可以重复几十到几百次,且形成反折式的二级结构。
2.端粒酶:一种RNA-蛋白质复合物,可以催化端粒不依赖模板的复制,从而补偿复制中引物去除导致的DNA分子的缩短。端粒的这种生长方式称为爬行模型(inchworn model)。
五.DNA甲基化
DNA忠实的复制也意味着亲代DNA中甲基化碱基的分布方式同样复制在子代DNA分子中。
原核生物DNA中含有一些甲基化的腺嘌呤和胞嘧啶,真核生物DNA含有一定量甲基化的胞嘧啶。甲基化碱基在DNA中的分布方式是特定的,这种特定的分布方式在真核生物中可能同细胞分化有关。
根据DNA半保留复制的机制,新生的子代DNA双链中的一条链来自亲代,因此是已甲基化的,另一条链是新合成的,还未甲基化,所以新生DNA是半甲基化的双链。
甲基转移酶以半甲基化DNA双链为底物,找到甲基化碱基的位置,并把新生链中相应位置的碱基予以甲基化。也存在不需要模板的甲基转移酶和脱甲基酶。染色体的结构可能决定它们的作用部位。
第四节 DNA损伤修复
一、突变
(一)突变的概念
突变(mutation [mju(?)"teiSEn])是指一个或多个脱氧核糖核苷酸的构成、复制或表型功能的异常变化。
又称DNA损伤(DNA damage),即遗传物质结构的改变引起遗传信息的改变。
(二)突变的意义
突变就其后果而言,并非都是危害生命的,也有积极意义,且在生物界普遍存在。
1.突变是进化、分化的分子基础。
自然界的生物进化过程是突变造成的。没有突变就没有遗传学。突变是缓慢而长期的。长期突变积累造成同一物种的个体差异。
自发突变或自然突变――不知道发生突变的真正原因。
2.只有基因型改变的突变、而没有可察觉的表型改变
例如:密码子第三碱基的改变、蛋白质非功能区编码基因序列的改变等。
DNA多态性(polymophism)是描述个体间的基因差别现象的。
多态性分析技术的应用:法医学的个体差别、亲子鉴定,临床医学上的器官移植配型,预防医学对疾病易感性分析等。
3.致死性的突变
突变发生在生命过程的重要基因。可导致个体和细胞死亡。人类利用此来消灭有害的病原菌。
4.突变是某些疾病的发病基础
遗传病、肿瘤、有遗传倾向的疾病(高血压、糖尿病、溃疡、肥胖等)。内科学的4000余种病,1/3以上属于遗传性或有遗传倾向的疾病。
人们认为突变有害,就是指这类突变。
二、引发突变的因素
自发突变的频率极低,在10-9左右。但是高等生物基因组庞大、细胞繁殖快,所以突变的作用不容忽视。
实验室用生活环境中导致突变的因素可以诱发突变。
1.物理因素:
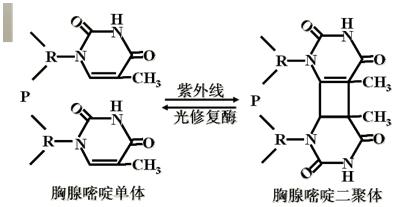
环境中的射线(紫外线和各种其它辐射),其中紫外线研究较多。
UV可以导致DNA分子上相邻的两个嘧啶碱基发生共价交联,生成嘧啶二聚体(环丁基环)。
2.化学因素:
很多的化学诱变剂同时就是致癌剂。包括:①碱基类似物:5-溴尿嘧啶(5-BU),②羟胺类(NH2OH);③亚硝酸盐类(NO2―);④烷化剂(氮芥类)。
来源:
(1)化工原料、化工产品、化工副产品
(2)工业排放物、汽车排放的废气
(3)农药、食品防腐剂或添加剂。
目前已经检出6万多种,而且每年增加超过千种。
物质和代谢过程中产生的自由基等因素能直接损伤DNA或干扰DNA的复制过程。
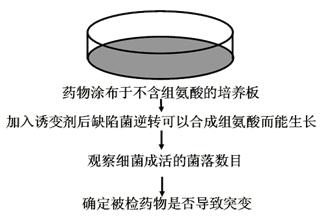
致癌物的检测方法
检测物质是否对细菌有突变作用(Bruce Amaes试验)。试验选用有缺陷的沙门菌:
(1)His―:组氨酸异养型(必须加入组氨酸才生长)。
(2)细胞壁缺陷:化学物质易透入。
(3)修复系统不活化(缺陷)。
三、突变分子改变的类型
(一)错配(点突变)
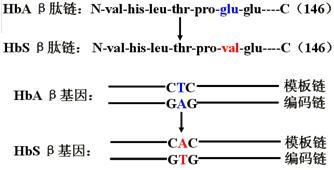
碱基的置换。发生在氨基酸的编码区,导致氨基酸的改变。
如:镰型红细胞贫血HbS = α2β26Glu→Val(β链的6位谷氨酸→缬氨酸)
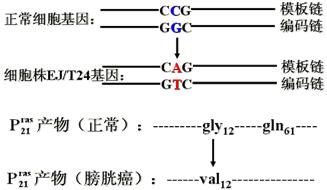
另外,膀胱癌细胞c-rasH基因点突变(GGC→GTC),导致表达产物12位Gly→Val。
(二)缺失、插入与框移突变
三联密码的改变,翻译出的蛋白质可能完全不同,缺失或插入点以后的密码全部改变.

(三)重排(重组)
DNA分子内发生较大片段的交换。交换(重排)后的两段基因都发生突变。
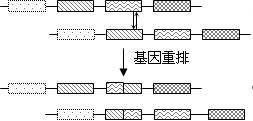
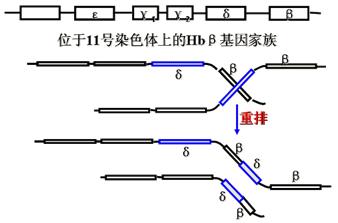
基因交换可以发生在一条分子内或两条分子间。下面是基因重排导致的两种地中海贫血。
四、DNA损伤的修复(DNA repairing)
未修复的损伤可能导致突变,降低细胞存活能力甚至致死。但正常的细胞具有多种途径用来除去损伤或不配对的核苷酸并代之以正确的核苷酸。
DNA复制中的错配可以通过“即时校读”加以纠正,此外还有修复机制。
(一)光修复(light repairing)―针对紫外损伤形成的嘧啶二聚体。
光修复酶(光复活酶)(photolyase)。
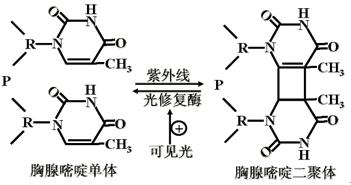
紫外线照射可能使相邻的胸腺嘧啶形成二聚体,有一种酶可分解这种二聚体,但需可见光(300~600nm),因此这个过程称为光活化反应。
如果二聚体还未被修复而复制叉已先通过了这一点,那么新生链上会留下一个缺口,在这种情况下,经DNA体内重组,受损伤的链仍可获得正确的模板用以修复。
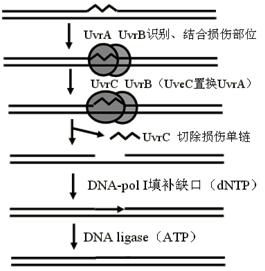
(二)切除修复――细胞内最重要的修复方式。
主要步骤(原核生物与真核生物的酶系统不同):
1.先把接近损伤部位5'端的磷酸二酯键切断,切除损伤部位及其邻近的核苷酸。
2.利用未受损的互补链作为模板,合成正确的新链。
3.缺口的连接
在某些情况下,糖苷酶能识别和水解受损碱基的糖苷链,然后插入酶把一个正确的碱基转移到空缺的糖基上去。
(三)重组修复
当DNA分子的损伤面积较大、来不及修复完善就进行复制时,损伤部位无模板功能,复制出来的子链回出现缺口。
1.将健康母链与缺口部分交换
重组蛋白(RecA、RecB、RecC等)参与。
健康母链:同一细胞内已经完成复制的链或来自亲代的一条链。
2.健康母链缺口的填补
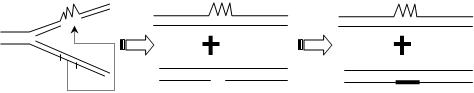
特点是:损伤部位未切除,仍然存在的损伤链随复制的次数增加而减少比例(被稀释)。
(四)SOS修复
是一种应急式的修复方式。在损伤严重、难以继续复制的情况下进行。
特点:特异性低、对碱基的识别、选择能力差,DNA保留的错误多,容易引起广泛、长期的突变。但经过此修复过程,复制可以继续、细胞可以存活。
第五节 逆转录现象和逆转录酶
一、逆转录酶
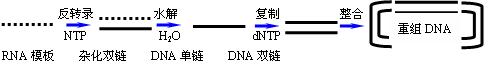
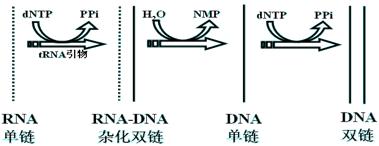
作用有三方面:
1.RNA作模板合成DNA单链,形成RNA-DNA杂化分子。
2.水解杂化分子中的RNA单链,生成DNA单链模板。
3.DNA作模板合成DNA单链,形成双链DNA分子。
2.存在于致癌的RNA病毒、蛙卵、白细胞、滋养层细胞。
二、逆转录酶的应用
病毒致癌理论研究。
制备基因工程中的目的基因(cDNA的制备)。