
第十四章 基因表达调控
第一节 基因表达调控的基本概念与原理
一、基因表达的概念和规律(特点)
(一)基因表达(gene expression)
1.基因表达的概念:
在一定调控机制控制下,基因经基因激活、转录、翻译等过程产生具有特异生物学功能的蛋白质,从而赋予细胞或个体一定的功能和遗传性状的过程。
简言之,基因表达就是基因转录及翻译的过程。
2.基因表达扩展:
并非所有的基因表达都产生蛋白质,有的基因表达只产生RNA。
所以编码rRNA和tRNA的基因转录过程也是基因表达。
(二)基因表达的规律
1.具有时间特异性或阶段特异性(temporal or stage specificity)
某一特定基因的表达严格按照特定的时间顺序发生。表现在:
(1)某一特定mRNA编码基因的表达随感染(病毒或细菌)的时间顺序发生。
随感染阶段的发展、环境的变化,有的基因开启、有的基因关闭,完全按功能需要。
(2)某一特定mRNA编码基因的表达随发育、生长(多细胞生物)的时间顺序发生。
多细胞生物基因的开启与关闭与从受精卵到组织器官发育形成的各个阶段保持一致。
多细胞基因表达的时间特异性又称为阶段特异性。
2.基因表达具有空间(细胞、组织)特异性(spatial or cell or tissue specificity)。
在个体生长全过程,某种基因产物在同一个体按不同组织空间顺序出现。即在空间分布上有差异。表现在:
(1)多细胞生物个体发育、生长的某一个阶段,同一基因表达产物在不同组织器官、组织空间表达量不同。
(2)多细胞生物个体发育、生长的某一个阶段,不同基因表达产物在不同的组织器官分布也不完全相同。
换言之,真核细胞中只有部分基因有转录活性,此部分基因在不同的组织、细胞的空间分布上有差异。在不同的细胞和同一细胞的不同发育阶段,基因表达的种类和数量不同。
基因表达的规律由特异基因的启动子(启动序列)和(或)增强子与调节蛋白之间的相互作用决定。
二、基因表达的方式或调节类型(不同基因对环境的反应性)
(一)基本表达(组成性表达)
1.管家基因(housekeeping gene):
某些基因在一个生物个体的几乎所有细胞中持续表达,称为管家基因。
该基因较少受环境因素的影响。
其产物对生命全过程都是必需的或必不可少的。
2.基本的或组成性(型)的基因表达(constitutive gene expression)
管家基因的表达方式称为基本的或组成性的基因表达。
3.管家基因表达的调控:
只受启动序列或启动子与RNA聚合酶相互作用的影响,不受其它机制调节。
(二)诱导及阻遏表达
1.适变基因
基因表达随外环境信号的变化而变化,此类基因称为适变基因。
2.适变基因的表达方式有2种
(1)可诱导基因的诱导
其中有些基因对环境信号应答时被激活、基因表达产物增加,该基因称可诱导基因。
可诱导基因在特定环境中表达增强的过程或表达方式称为诱导(induction)。
如:DNA损伤→修复酶基因被诱导激活→修复酶增加。此类基因多数是参加分解代谢的酶类。
(2)可阻遏基因的阻遏
其中有些基因对环境信号应答时被抑制、基因表达产物降低,该基因称可阻遏基因。
可阻遏基因表达产物水平降低的过程或表达方式表达方式称为阻遏(repression)。
如:色氨酸→编码色氨酸合成酶的基因表达被抑制→色氨酸合成减少。此类基因多数是参加合成代谢的酶类。
3.适变基因表达的调控
除了受启动序列或启动子与RNA聚合酶相互作用的影响,还受其它机制调节。
一般此类基因的调控序列中含有特异刺激的反应元件。
(三)协调表达与协调调节
1.协调表达和协调调节(coordinate expression and coordinate ragulation)
在一定机制控制下,功能相关的一组基因,无论何种表达方式,均需要协调一致、共同表达,即是协调表达。该调节称为协调调节。
2.协调调节的意义
可以使参与同一代谢途径的蛋白质(包括酶和转运蛋白及其它蛋白因子)分子比例适当,确保代谢途径有条不紊地进行,从而适应环境。
三、基因表达调控的生物学意义
(一)适应环境、维持生长和繁殖
1.原核生物、单细胞生物:
调节基因的表达是为了适应环境、维持生长和细胞分裂而表达。如:细菌乳糖操纵子的调节。
2.高等生物也普遍存在适应性表达方式。
如:长期酗酒者体内醇氧化酶(脱氢酶)基因表达增强。
(二)维持个体发育和分化
多细胞生物个体生长发育的不同阶段、同一阶段的不同组织器官,蛋白质的种类、含量存在很大的差异。这些差异是调节细胞表型的关键。
高等哺乳类动物的各种组织器官的分化、个体发育都由特定的基因控制。
四、基因表达调控的基本原理
(一)基因表达的多级调控
基因表达需要细胞的严密调控机制,并且是多级水平的调控,可以发生在遗传信息传递的各个环节。包括:基因激活、转录起始、转录延长、转录后加工、mRNA降解、蛋白质翻译、翻译后加工修饰、蛋白质降解等多级水平进行。
但以转录水平的调控最重要。基因转录起始或激活是基因表达的基本控制点。
(二)基因转录激活调节的基本要素
1.特异DNA序列――基因的结构
(1)原核生物细胞――操纵子机制
(2)真核生物细胞――顺式作用元件(编码基因两侧的DNA序列)
在分子遗传学中:
顺式(cis-)作用指相对于同一分子或染色体而言;即:分子内作用。反式(trans-)作用指相对于不同的分子或染色体而言。即:分子间作用。
顺式作用元件(cis-acting element):
可影响自身基因表达的DNA序列。通常是非编码序列。如增强子对启动子的作用。不同的真核生物基因中常含有一些共有序列(核心序列:TATA盒、CCAAT盒等):启动子、增强子、沉寂子等。顺式作用元件并非都位于转录起始点上游(如增强子)。
2.调节蛋白
(1)原核生物基因的调节蛋白都属于DNA结合蛋白,分三类:
1)特异因子:决定RNA聚合酶对一个(或一套)启动序列的特异性识别、结合能力(如:σ)。
2)阻遏蛋白:可以结合特异的DNA序列――操纵序列,阻遏基因转录。介导负性调节。
3)激活蛋白:可以结合启动序列邻近的DNA序列,促进RNA聚合酶与启动序列结合,增强RNA聚合酶活性。介导正性调节。如:分解(代谢)物基因激活蛋白(CAP)。
激活蛋白有时对某些基因是必需的。无激活蛋白时RNA-pol很少或不能结合启动部位。
(2)真核生物基因调节蛋白
又称转录因子(transcription factor)或反式作用因子(trans-acting factor)
1)反式作用与反式作用因子:
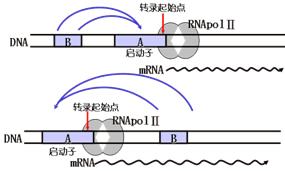
绝大多数真核转录调节因子是DNA结合蛋白,由某一个基因表达后(蛋白质)特异地与另一个基因的顺式作用元件(DNA)作用,反式激活该基因的转录。如下图。

2)顺式作用与顺式作用蛋白
有些基因产物可以特异地识别、结合自身基因的调节序列,调节自身基因的开启与关闭。这就是顺式作用。具有这种调节方式的调节蛋白称顺式作用蛋白。
3.DNA-蛋白质、蛋白质-蛋白质相互作用
(1)DNA-蛋白质相互作用
指反式作用因子与顺式作用元件之间的特异识别与结合。通常是非共价键的结合。
被调节蛋白识别并结合的DNA位点通常呈对称或不完全对称结构。
(2)蛋白质-蛋白质相互作用――形成二聚体或多聚体
1)二聚化(dimerization):
两分子单体通过一定的结构域结合成二聚体。是调节蛋白结合DNA时最常见的形式。
同种分子形成同二聚体(homodimer),异种分子形成同二聚体(hEterodimer)。异二聚体DNA结合能力更强。但有时调节蛋白二聚化后可能丧失结合DNA的能力。
2)有些调节蛋白不能直接结合DNA
通过蛋白质-蛋白质相互作用参与DNA-蛋白质复合物的形成,间接结合DNA,调节基因转录。在真核生物常见。
4.RNA聚合酶活性――受启动序列/启动子结构及其调节蛋白的性质影响。
(1)启动序列/启动子与RNA聚合酶活性
1)原核启动序列、真核启动子的组成包括:
转录起始点、RNA-pol结合点、控制转录的调节组件。启动子比启动序列结构复杂。
上述核苷酸序列影响其与RNA聚合酶的亲和力。同样影响转录启动的频率。
2)真核RNA聚合酶单独存在时与启动子的亲和力极低或没有亲和力。
必须与基本转录因子形成复合物后才能与启动子结合。
(2)调节蛋白与RNA聚合酶活性
1)环境信号→调节蛋白表达→DNA-蛋白质、蛋白质-蛋白质作用→RNA聚合酶→表达。
2)热休克反应(heat shock response):
原核特异因子σ可以改变RNA聚合酶识别启动序列的特异性。
细菌热应激时,RNA-pol全酶中的σ70被σ32所取代,这时RNA-pol会由识别常规启动序列转向识别并结合另外一套启动序列,启动一套基因表达,即所谓的热休克反应。
真核细胞普遍存在的热休克基因需要热休克蛋白(Hsp)才能启动,Hsp与原核的σ32在氨基酸组成和排列上高度同源。
第二节 原核基因转录调节
一、原核基因转录调节的特点
(一)σ因子决定RNA聚合酶识别特异性
原核生物细胞仅有一种RNA聚合酶,全酶参与转录的起始,其中不同的σ因子(亚基)识别特异的启动序列,决定特异基因的转录激活。核心酶参与转录的延长。
(二)操纵子模型的普遍性(大多数原核基因表达的调控模式)
操纵子(元)(opron):
原核生物的几个功能相关的结构基因往往排列在一起,在同一调控机制下,转录生成一段mRNA,然后分别翻译成几种不同的蛋白质(催化某代谢过程的酶或共同完成某种功能)。
这些结构基因与上游的启动子、操纵基因共同构成转录单位,称操纵子(元)。
(三)阻遏蛋白与阻遏机制的普遍性
原核基因调控普遍涉及特异阻遏蛋白参与的开、关调节机制。特异的阻遏蛋白是控制原核生物启动序列活性的重要因素。
二、乳糖操纵子的调节机制
(一)一般操纵子的结构
操纵子由一组结构基因和其上游的启动序列及操纵基因组成。在调控区上游一般还有编码阻遏蛋白的阻遏物基因。结构基因是蛋白质的编码系列。启动序列是RNA聚合酶的结合部位,其中含有两个一致性序列:①―10bp附近的TATAAT②―35bp附近的TTGACA。操纵基因是结构基因和启动序列之间的阻遏蛋白的结合部位和RNA-pol通过的开关。
根据阻遏蛋白与小分子物质作用性质不同可把操纵子分为可诱导型和可阻遏型。
(二)阻遏蛋白的负调控与可诱导型操纵子
1.负调控的概念:
原核生物基因的表达一般受蛋白质(阻遏蛋白)的抑制,阻遏蛋白与操纵基因结合后可抑制基因的转录。介导的是负性调节。
阻遏蛋白与操纵基因的结合受一些小分子物质的调节。
2.可诱导型操纵子:
其结构基因一般是参与分解代谢的酶类。如乳糖操纵子(E.coli的lac操纵子)。
其结构如下:
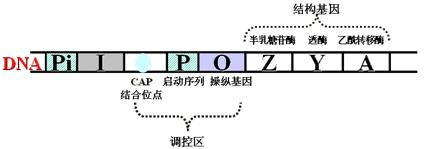
(1)三个结构基因:Z、Y、A基因。编码利用乳糖的三种酶(β-半乳糖苷酶、透酶、乙酰转移酶)。
(2)一个操纵基因:O基因,可以和阻遏蛋白结合,控制结构基因的转录(负性调节)。
(3)一个启动序列:P序列,可和RNA聚合酶结合。此位点是启动子部位。
(4)一个调节基因:I基因,编码阻遏蛋白。
(5)分解代谢基因活化蛋白(CAP)结合位点:结合CAP-cAMP复合物。
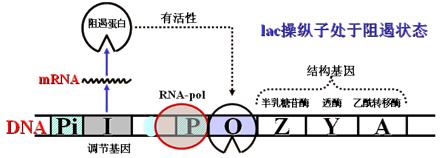
1)在无诱导物(代谢底物如乳糖)时:
P1启动序列操纵表达的阻遏蛋白与操纵基因结合,使其处于关闭状态,RNA-pol不能通过,基因不表达。(阻遏并非绝对,只是表达量很少,只有寥寥的β-半乳糖苷酶和透酶)。
2)有诱导物存在时:
阻遏蛋白与操纵基因分离(如:诱导物半乳糖可与阻遏蛋白结合使其失去结合操纵基因的活性),操纵基因处于开放状态,RNA-pol能通过,基因可以表达,表达产物的酶类对底物进行分解代谢。

注意:
真正的诱导剂是乳糖经透酶作用进入细胞后由β-半乳糖苷酶催化转变成的半乳糖。
异丙基硫代半乳糖苷(IPTG)是实验室常用的强的诱导剂,不被细菌代谢而十分稳定。
(三)CAP介导的正性调节与cAMP
1.正调控的概念:
一些蛋白质如分解代谢基因活化(激活)蛋白(CAP)与操纵子DNA结合后,可促进转录过程。CAP属于同二聚体,分子内含有DNA结合区以及cAMP结合位点。
当有cAMP存在时与CAP结合,使后者构象改变并与CAP结合位点结合,刺激RNA转录活性,介导正性调节。在原核生物中正调控方式很少见。
2.cAMP对乳糖操纵子转录的调控:
乳糖操纵子的RNA聚合酶结合部位(P)上游附近有一个CAP的结合位点。但CAP须与cAMP结合后才能与该部位结合。cAMP通过作用于CAP发挥对转录的调节作用属于正调控。
(1)当培养基中无葡萄糖存在、细胞内cAMP水平高时:
CAP与cAMP结合形成cAMP―CAP复合物,然后与CAP结合部位结合,促进RNA聚合酶与启动子结合和进行转录,产生代谢乳糖酶类。
(2)当培养基中有葡萄糖存在、细胞内cAMP水平低时:
cAMP―CAP复合物不能形成,游离的CAP不能与CAP结合部位结合,RNA聚合酶则不能与启动子结合和进行转录。
这使得葡萄糖与乳糖同时存在时,葡萄糖可被优先利用。消耗完葡萄糖后才利用乳糖。
(四)协调调节
1.阻遏蛋白的负性调节与CAP的正性调节协调
(1)当lac阻遏蛋白封闭转录时,CAP对系统不能发挥作用
(2)如果没有CAP加强转录活性,即使阻遏蛋白从操纵基因上解聚仍没有转录活性。
两种机制相辅相成、互相协调、互相制约。二者缺一不可。
2.葡萄糖的分解代谢阻遏
(1)当葡萄糖和乳糖共同存在时,细菌首先利用葡萄糖才是最节能的。
葡萄糖通过降低cAMP浓度阻碍cAMP与CAP结合而抑制lac操纵子转录,称分解代谢阻遏(catabolic repression)。
(2)lac操纵子强的诱导作用的条件是:乳糖存在的同时又要缺乏葡萄糖。
三、其它转录调节机制
(一)可阻遏型操纵子(色氨酸操纵子)――转录衰减
其结构基因表达产物一般是参与某种物质合成代谢的酶类。如色氨酸操纵子。
1.在细胞中合成产物(色氨酸)丰富时:
合成产物作为(辅)阻遏物与辅阻遏蛋白结合,使后者有活性,能与操纵基因结合操纵基因则处于关闭状态,基因不表达。
如色氨酸与辅阻遏蛋白结合,形成能与操纵基因结合的活性阻遏蛋白。色氨酸称为辅阻遏剂或辅阻遏物。
2.在合成产物(色氨酸)不足时:
(辅)阻遏蛋白不与操纵基因结合,操纵基因处于开放状态,基因表达,基因表达产生的酶可对产物进行合成以补充其不足。
随色氨酸含量的升高,此后的转录速率受转录衰减(attenuation)机制的调节。
(1)trp操纵子的衰减子区域(attenator region):
色氨酸操纵子上位于第一个结构基因与启动序列(P)之间的区域。
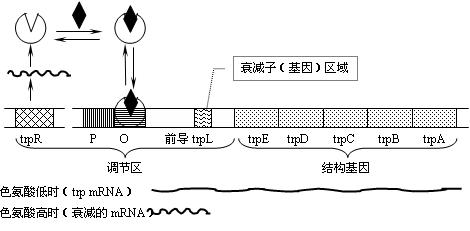
(2)衰减子(attenutor):
mRNA分子上的不依赖ρ因子(Rho)的转录终止结构(由操纵子上的衰减子区域编码)。
可以使RNA聚合酶脱落而终止转录。
(3)转录衰减机制――转录与翻译的偶联进行导致衰减子的形成
色氨酸操纵子的mRNA序列1中有两个色氨酸密码子,翻译时如果色氨酸浓度很高,则核蛋白体很容易通过序列1,并且封闭序列2。
色氨酸操纵子的mRNA序列3和序列4形成衰减子结构,使前方的RNA聚合酶脱落,转录终止。
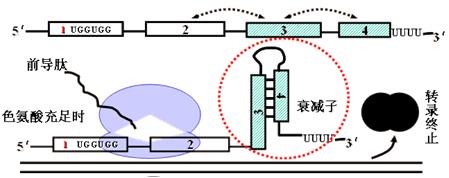
反之,如果色氨酸缺乏,则核蛋白体停止于序列1的色氨酸密码子前,序列2和序列3形成发夹结构,阻止了序列3和4形成衰减子结构,RNA聚合酶继续向前转录。
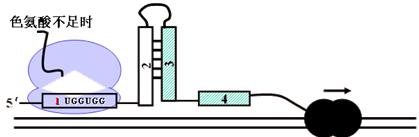
注意:
前述色氨酸缺乏并非绝对没有;不至于连酶合成的原料都没有;核蛋白体“停止”不如说是“停顿”,只是缓慢通过。
(二)基因重组(genetic recombination)
可以改变启动序列方向,开启或关闭某些基因的表达。下图为沙门菌鞭毛素基因表达的调节。

(三)原核生物转录的整体调控模式与SOS反应
1.调节子:
是由成群操纵子组成的高一级调控网络,这些操纵子受共同调节蛋白的调控,以适应内外环境的变化。它属于原核生物整体调控方式。
2.DNA损伤修复的SOS系统:
(1)SOS反应(SOA response):
细菌(E.coli)经紫外线照射发生DNA损伤或复制抑制而诱发一系列表型变化的现象或过程。SOS反应是调节子模式的典型例子。
(2)SOS反应机制
参与SOS修复的酶和蛋白质如RecA、RecC、UvrA等的基因(SOS基因)分散在染色体的不同操纵子中,而在基础状态时这些操纵子由一个共同的阻遏蛋白LexA抑制。
在紫外线等外界因索作用下,DNA损伤产生大量的单链缺口,损伤的DNA使细菌内RecA蛋白产生水解酶活性而水解阻遏蛋白(LexA),使之失去阻遏作用,从而使处于不同操纵子的recA、recC、uvrA等基因表达表达出RecA、RecC、UvrA,参与DNA修复。
第三节 真核基因转录调节
一、真核生物基因的特点
(一)真核生物基因组结构庞大(30亿碱基对)――分三种类型
1、4万以上的基因(按每个基因1500bp计,约占全基因组的6%),编码5千-1万种mRNA。清楚的仅有几千基因,已有40人获奖。
2、重复序列占5-10%(编码rRNA等)。
3、没有直接的遗传学功能的基因占80-90%(jun基因)。且DNA在细胞核内,与组蛋白构成以核小体为单位的染色质,染色质结构影响基因转录。除配子细胞外的体细胞都是二倍体(两套相同的基因组)。基因表达调控机制复杂。
(二)以单个基因为转录单位,转录产物为单顺反子。
单顺反子即一个编码基因转录成一个mRNA分子、翻译成一条肽链。即使两个基因相邻很近也分别转录。
(三)基因组往往存在着许多重复序列。有反转重复序列。
1.重复序列长短不一,从10个核苷酸以下到数百、数千不等。
2.重复的频率不同:
(1)高度重复序列:出现106次(百万次)。人10-20%。卫星DNA属于此列。有些A、T序列丰富,易于解链。
(2)中度重复序列:103~104次(千次~万次)。人20-30%。
(3)低度重复序列:低于千次。
(4)单拷贝序列:只出现一次或很少出现。人60-65%。除rRNA、tRNA、组蛋白和免疫球蛋白、糖代谢的酶等少数基因外,多数真核基因都是单拷贝基因。
3.反转重复序列:两个互补序列在同一DNA链上反向排列。
(四)真核基因是不连续的,多为断裂基因,编码序列被内含子间隔。内含子功能不清,可能参与调控。
二、真核基因表达调控特点(与原核不同)
(一)RNA聚合酶
三种真核RNA聚合酶分别转录三种RNA。单独没有活性,必须和转录因子共同作用。每种RNA聚合酶包括约10个亚基,有些是共有的(相同),有些是每一种酶特有的。
其中TATA盒结合蛋白(TBP)是三种酶共有的。RNA-polⅡ所需的TFIID是TBP和TBP相关因子(TAF)组成的蛋白复合物。TAF对于传递上游激活序列的信息十分重要。
(二)活性染色体结构变化:基因被激活后,染色体的相应区域发生结构和性质的变化:
1.对核酸酶极度敏感:超敏位点多在调节蛋白结合位点附近。
2.DNA拓扑结构变化:RNA聚合酶前方转录区为正超螺旋,后面的DNA则是负超螺旋。
3.DNA碱基修饰变化:活化后甲基化程度降低。基因表达的程度与甲基化的范围或程度是成反比的。活化前DNA约有5%的胞嘧啶被甲基化修饰,多发生在5/侧翼区的CpG序列(CpG岛)。
4.组蛋白变化
(1)富含赖氨酸的组蛋白(H1样组蛋白)水平降低。
(2)H2A?H2B二聚体稳定性降低。易于被从核心中置换出来。
(3)组蛋白被修饰(乙酰化、泛素化),使核小体结构不稳定。
(4)H3组蛋白的巯基暴露。
(三)正性调节占主导
由真核基因特点决定,采用正性调节更有效并且比负性调节更经济。
(四)转录与翻译不在同一个区域、也不能同时进行
先在细胞核转录,并且需要转录后加工;然后在细胞浆翻译。
(五)转录后的修饰、加工比原核细胞更加复杂
三、真核基因转录激活
(一)顺式作用元件的种类(启动子、增强子、沉默子等)
按功能特性划分。
1.启动子promotor:
是原核基因操纵子中启动序列的同义语。启动子是真核基因RNA聚合酶结合位点周围的一组转录控制组件(module),每组含7-20bp的DNA序列。
2.启动子的组成结构:
一个转录起始点(transcription start site ,initiation site)和一个以上的功能组件。转录起始点有时与功能组件无法区分。
(1)典型的功能组件:
TATA盒(共有序列:-TATAAAA-),通常的位置:―25至 ―30bp。
作用是基本转录因子TFⅡD的结合位点,控制转录起始的准确性和频率。
(2)常见的功能组件:
GC盒(GGGCGG)盒、CAAT(GCCAAT),位置:―30至 ―110bp
3.启动子的种类(三类)
(1)最简单的启动子:TATA盒 + 转录起始点
(2)典型的启动子:以最简单的启动子为基础,连同其上游的GC盒和(或)CAAT盒
(3)不含TATA盒的启动子(富含GC的和没有GC富含区的2种)
富含GC的启动子最初发现于管家基因,一般含有数个分离的转录起始点。
没有GC富含区的启动子有一个或多个转录起始点,多数活性很低或没有转录活性。
2.增强子(enhancer):
(1)概念:远离转录起始点(1~30kb)、决定基因的时间和空间特异性表达、增强启动子转录活性的DNA序列。
(2)作用特点:
1)作用与位置无关。可以在基因前面、后面、基因中,可在远距离影响启动子,与蛋白因子结合后对转录具有增强作用。
2)决定基因表达的时间、空间特异性。长度约200bp。
3)其发挥作用的方式与其与启动子的方向无关,可在3/一5/或5/一3/方向影响启动子。
4)增强子对启动子无严格专一性。或者无基因特异性。
5)没有增强子的启动子通常没有活性;不通过启动子,增强子无从发挥作用。
(3)增强子的组成(增强体):
增强子由若干增强体(enhanson)组成,有些增强体可以在启动子和增强子中都出现。
注意:
增强子和启动子经常交错覆盖或者连续,有时无法区分则统称启动子。文献中指的有独立转录活性和决定基因时间、空间特异性表达能力的启动子都属于此类。
另外,酵母基因中的上游激活序列(UASs)在转录激活中的作用及方式类似增强子。
3沉默子(silencer):
某些基因的负性调节元件。与特异性蛋白因子结合以后对基因转录起阻遏作用。
另外,在结构基因后面有加尾信号及其转录终止信号,虽然与转录激活无关,但与转录终止有关。常在polyA位点上游10-20bp,常见的序列是-AATAA-。去掉此序列则转录不停止。
(二)反式作用因子
1.反式作用因子:真核的转录调节因子称转录因子(transcription factor ,TF)。
是与顺式作用元件直接或间接作用的、由其它基因表达产生的调节蛋白。它们通过DNA―蛋白质或蛋白质―蛋白质之间的相互作用影响RNA聚合酶活性而调节基因的转录。
2.转录调节因子的分类:转录因子按功能特性分为两类
(1)基本转录因子(general transcription factor)
是RNA聚合酶结合启动子所必须的一组蛋白因子。决定三种RNA转录的类别。
不同的聚合酶有不同的基本转录因子,仅个别的基本转录因子可以通用(如:TFⅡD,其组成成分TBP本身就是酶的亚基),其它如TFⅡA、B、E、F、H等,都是RNA聚合酶Ⅱ转录mRNA所特有的。
(2)特异转录因子(special transcription factor)
是个别基因转录所必需的,决定该基因的时间、空间特异性表达。此类因子对基因转录的作用可以是激活,也可以是抑制。
1)转录激活因子:多数是增强子结合蛋白(EBP)。
2)转录抑制因子:多数是沉默子结合蛋白。有的通过蛋白质-蛋白质相互作用“中和”转录激活因子(EBP)或TFIID,降低其细胞内有效浓度,抑制转录。
3.转录调节因子的结构
(1)转录因子至少含有两个不同的结构域:
1)DNA结合域(DNA binding domain)

通常由60-100个氨基酸残基组成。最常见的结构形式是锌指(zinc finger)、碱性氨基酸组成的α螺旋、亮氨酸拉链(basic leucine zipper,bZIP)、碱性螺旋-环-螺旋(basic helix-loop-helix,bHLH)结构的碱性氨基酸伸展。
2)转录活化域(激活域)(actvation domain)。30~100氨基酸残基组成。根据氨基酸组成特点分为:
酸性激活域(acidic actvation domain)、谷氨酰胺富含域(glutamine-rich domain)、脯氨酸富含域(proline-rich domain)。
(2)其它蛋白结合域
很多转录因子具有介导蛋白质―蛋白质相互作用的结构域(其它蛋白结合域),最常见的是二聚化结构域。
在不同结构域中往往存在着在氨基酸组成和空间结构上具有一定规律的一致性结构,即基元。如螺旋―转角―螺旋(或螺旋―环―螺旋)、亮氨酸拉链和锌指结构等。这些结构是反式作用因子与DNA双螺旋结合及相互作用的空间构象。
(三)mRNA转录激活及其调节(参见第十二章 转录)
1.转录因子Ⅱ(TFⅡ)族:
是与RNA聚合酶Ⅱ(polⅡ)一起形成转录前起始复合物(PIC)的一类反式作用因子。RNA聚合酶Ⅱ的功能是转录mRNA,但其不能单独识别结合启动子。需要依赖基本转录因子和特异转录激活因子的存在。
2.转录前起始复合物(PIC)的形成:
必须由基本转录因子TFⅡD的组成成分TBP识别TATA盒或启动元件,并有TAF参与形成TFⅡD-启动子复合物,继而在TFⅡA-F参与下,RNA聚合酶Ⅱ、TFⅡD、TFⅡB形成前起始复合物(PIC)。PIC的装配过程如下:
TATA盒→TFⅡD→TFⅡA→TFⅡB→RNA-polⅡ/TFⅡF→TFⅡE→PIC
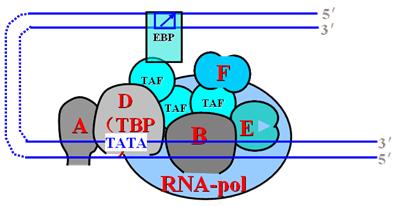
以上转录因子有不同作用。所有转录因子中只有TFⅡD直接与启动子中的TATA盒结合。TFⅡA可稳定TFⅡD与DNA的结合,TFⅡB的结合是关健步骤,可促进RNA-polⅡ的进入,TFⅡE和TFⅡF分别具有ATP酶和解旋酶活性,共同促进单链DNA模板的暴露。PIC形成后开始mRNA的转录。
3.稳定的转录起始复合物的形成
前转录起始复合物不稳定,还不能有效地启动转录。此时结合了增强子的转录激活因子(增强子结合因子:EBP)接近TFⅡD或通过TAF与TFⅡD联系,形成稳定的转录起始复合物。
基因转录激活过程就是形成稳定的转录起始复合物的过程。此时TBP相关因子与转录激活因子共同决定阶段特异性及组织特异性转录。