第五章 糖类代谢
第二篇 物质代谢及其调节
本篇重点:
基本反应途径、关键酶及其主要调节环节、重要的生理意义、各种物质代谢的相互联系、代谢异常与疾病的关系。
物质代谢及能量代谢如下图:

第五章 糖代谢
糖是自然界存在的一大类具有广谱化学结构和生物功能的有机化合物。它主要是由绿色植物经光合作用形成的。
糖是多羟基的醛或酮的统称。实验式Cn(H2O)m。这类物质主要由碳、氢和氧所组成,其分子式通常以Cn(H2O)m表示。由于一些糖分子中氢和氧原子数之比往往是2:1,刚好与水分子中氢、氧原子数的比例相同,过去误认为此类物质是碳与水的化合物。故有“碳水化合物(carbohydrate)”之称。
实际上有些糖(鼠李糖和脱氧核糖)中氢、氧原子数之比并非2:1。而一些非糖物质(甲醛、乳酸和乙酸)中氢、氧原子数之比却都是2:1,所以,称糖为“碳水化合物”并不恰当。
糖是生物界中分布极广、含量最丰富的一类有机物质,几乎所有的动物、植物、微生物体内都含有它。其中以植物界最多,约为85―95%(占其干重的80%)。
生物细胞内、血液里也有葡萄糖或由葡萄糖等单糖物质组成的多糖(如肝糖原、肌糖原)存在。人和动物的器官组织中含糖量不超过组织干重的2%,微生物体含糖量约占菌体干重的10―30%,糖与蛋白质、脂类结合成复合糖存在。
糖类物质的生物学作用是为生命活动提供能源和碳源。主要是通过氧化而放出大量的能量,以满足生命活动的需要。人类能量的50-70%来自糖(主要是葡萄糖)。
淀粉、糖原是重要生物能源,它也能转化为生命必需的其它物质,如蛋白质和脂类物质。纤维素是植物的结构糖。
分类:
糖类物质以它们水解的情况分类:单糖、寡糖、多糖和结合糖(糖复合物)
1、凡不能被水解成更小分子的为单糖。单糖又可根据糖分子含碳原子数多少分类。
在自然界分布广、意义大的是五碳糖和六碳糖,它们分别称为戊糖和已糖。核糖、脱氧核糖属戊糖,葡萄糖、果糖和半乳糖为己糖。
2、凡能水解成少数(2―6个)单糖分子的称为寡糖,其中以双糖存在最为广泛。蔗糖、麦芽糖和乳糖是其重要代表。
3、凡能水解为多个单糖分子的糖为多糖,其中以淀粉、糖原和纤维素等最为重要。
4、与非糖物质结合的糖称复合糖,加糖蛋白和糖脂。
5、糖的衍生物称衍生糖,如糖胺、糖酸和糖酯等。
单糖的种类很多,以葡萄糖(游离的,结合形式的)数量最多,在自然界分布亦广。
单糖的结构及性质各有异,但相同之处也不少。葡萄糖结构及性质有代表性。
葡萄糖可以游离存在于水果、谷类、蔬菜和血液中,也可以结合形式存在于麦芽糖、蔗糖、淀粉、糖原、纤维素及其它葡萄糖衍生物中。
一、单糖
丙糖:最简单的单糖。
丙糖中的醛糖是甘油醛,有一个不对称碳原子,构型有D-甘油醛和L-甘油醛。
凡是可视为D-甘油醛衍生物的糖都是D糖、视为L-甘油醛衍生物的糖都是L糖。
丙糖中的酮糖为二羟基丙酮。
自然界中的单糖多为醛糖,以己糖最普遍和重要,戊糖次之。
常见的单糖如下表:
| 醛糖 | 酮糖 | |
| 丙糖(3C) | D-甘油醛 | 二羟丙酮 |
| 丁糖(4C) | D-赤藓糖 | |
| 戊糖(5C) | D-核糖 D-2-脱氧核糖 |
D-木酮糖 D-核酮糖 |
| 己糖(6C) | D-半乳糖 D-甘露糖 D-葡萄糖 |
D-果糖 |
| 庚糖(7C) | D-景天庚酮糖 | |
| 辛糖(8C) | D-辛酮糖 |
己醛糖(己醛糖中以葡萄糖分布最广):是构成淀粉、糖原、纤维素及其他许多糖类物质的基本单位。葡萄糖是人类血液中的正常成分,给机体提供能量的重要物质。
己糖多以比较稳定的1:5氧桥(1氧:5碳)的六元环结构(吡喃型己糖)存在。
吡喃型已糖常与极少量1:4氧桥(1氧:4碳)五元环结构糖(呋喃型己糖)成平衡状态。
己糖透视式中:
D、L构型的确定是以C5上羟甲基在含氧环上的排步确定,α、β构型的确定是以C1上的半缩醛羟基在含氧环上的排布而决定的:
氧环上的碳原子按顺时针排列,(C5)羟甲基在平面之上为D-型,在平面之下为L-型。
在D-型中,半缩醛(C1)的羟基在平面之下为α型,在平面之上为β型。
葡萄糖的结构与构型

在溶液中:
α-D(十)-葡萄糖和β-D(十)-葡萄糖分别占总碳含量的36和63%,而链式结构葡萄糖仅占总碳含量的1%以下。
构型(configuration)
是指一个分子由于其中各原子特有的固定的空间排列,而使该分子所具有的特定的立体化学结构。当某一物质由一种构型转变为另一种构型时,要求共价键的断裂和重新形成。
不对称碳原子是指连接四个不同原子或基团的碳原子。
镜象对映体
互为镜象不能重叠的两类化食物称为一对“对映体(antipode)”,其两种构型,一个为D-型,一个为L-型。
旋光异构现象和旋光度
当光波通过尼克棱镜(nicol prism)时,由于棱镜的结构通过的只是沿某一平面振动的光波,其他都被遮断,这种光称为“平面偏振光”。
当它通过具有旋光性质的某异构物溶液时,则偏振面会向右(顺时针方向)或向左(反时针方向)旋转。使偏振光振动面右旋的称右旋光性物质,而使偏振光振动面左旋的称左旋光性物质。用“+”和“―”右旋和左旋。
旋光度是旋光性物质的一种物理性质。它和其他物理性质一样,在一定的条件下是一个常数。为了能使各种旋光性物质的旋光度均为常数必须把影响它们旋光度的因素如温度、浓度、光的波长、旋光管的长度等加以固定。
D、L是指构型,“十”、“―”指旋光方向。D与“十”、L与“―”并无必然联系。
D-葡萄糖和D-果糖旋光方向分别为“十”和“―”,而L-葡萄糖与L-果糖旋光方向却均为“―”。构型与旋光方向是两种概念。
二、寡糖
寡糖中最重要的是双糖:
其中常见的是蔗糖(葡萄糖+果糖)、乳糖(葡萄糖+半乳糖)和麦芽糖(葡萄糖+葡萄糖)。
三、重要的多糖
1、淀粉:
是植物贮存的养料,是人类食物中的主要成分,有直链、支链之分.
葡萄糖分子间多是α(1→4)糖苷键,而其分支点上则是α(1→6)糖苷键。
能为酸或α淀粉酶所水解。
2、糖原(动物淀粉):
是动物组织中储存的养分,其分子量较淀粉的略大,分支较支链淀粉略多,性质与淀粉的大致相似。
3、纤维素:
是构成植物躯干的主要成分,它由许多β-D-葡萄糖分子通过β(1→4)糖苷键缩合生成。
第一节 概述
一、糖的生理功能
(一)提供能量是糖类的最主要的生理功能:
人体所需能量的50―70%来自糖。16.7kJ/g。
(二)糖的中间产物可转变为其它含碳化合物:
氨基酸、脂肪酸、核苷(酸)等。
(三)衍生为生理活性物质:
辅酶(NAD+、FAD)、核(苷)酸、ATP等。
(四)糖是组成人体结构的重要成分:
结缔组织、软骨、骨基质、各种膜结构成分。
(五)参与细胞识别、信息传递、免疫功能。
二、糖的消化
人类食物中的糖:
植物淀粉和动物糖原以及麦芽糖、蔗糖、乳糖、葡萄糖等是主要的糖。
其中,植物光合作用产生的淀粉是动物的重要营养来源。
食物中的大量纤维素因人体不含有β-糖苷酶而无法利用。其作用是促进肠蠕动,也是维持健康所必需。
(一)消化部位
1、口腔:
唾液淀粉酶水解淀粉的α-1,4-糖苷键。胃酸使该酶失活。
(胃中不含有消化糖类的酶)。
2、小肠――消化糖类的主要场所。
(1)肠液消化:
胰α-淀粉酶,水解淀粉的α-1,4-糖苷键。产物有二糖、三糖、寡糖。
(2)接触消化:
微绒毛表面吸附的胰淀粉酶,继续水解淀粉和寡糖。
(3)膜消化:
微绒毛中的酶(质膜成分)。产物是单糖。
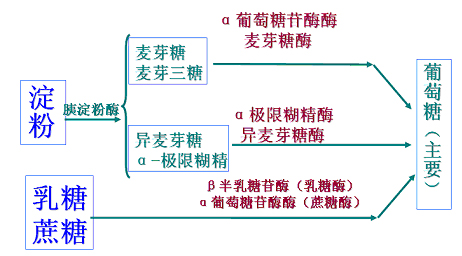
此类酶包括:
α糊精酶、糖淀粉酶、麦芽糖酶、异麦芽糖酶、乳糖酶、蔗糖酶。
(二)消化过程
二糖及寡糖经小肠上皮细胞分泌的寡糖酶(oligosaecharidase)从非还原末端水解。
α-葡萄糖苷酶(α-glucosidase)(包括麦芽糖酶)水解没有分支的麦芽糖和麦芽三糖;
α-极限糊精酶(包括异麦芽糖酶)则水解α-极限糊精和异麦芽糖的α1,4-和α1,6-糖苷键,生成葡萄糖。
蔗糖由α-葡萄糖苷酶(α-glucosidase)水解,而乳糖则由β-半乳糖苷酶(β-glactosidase)分解。(细胞中也有蔗糖酶和乳糖酶)。
最终消化产物绝大多数是单糖。
经胰α-淀粉酶(α-amylase)水解淀粉产生:
1、二糖:
麦芽糖(mactose)(葡萄糖α-1,4-葡萄糖)――40%
异麦芽糖(葡萄糖α-1,6-葡萄糖)――(5%)
2、三糖:
麦芽三糖(α-1,4-三聚葡萄糖)――(25%)
3、寡糖:
麦芽寡糖(α-1,4-四聚糖~五聚糖)少量
α-极限糊精(临界糊精、支链寡糖,4―9碳糖)――(30%)
肠液消化的酶、过程及产物
三、糖的转运与吸收
食物中的糖经消化后的单糖:被小肠粘膜细胞吸收进入血液,经门静脉入肝脏。
D-葡萄糖、D-果糖、D-半乳糖等。
未被消化的二糖、寡糖及多糖:不能被吸收。
它们经肠道细菌的分解后以酸、CH4、CO2或H2的形式放出或参加代谢。
注:糖的消化与吸收是同步进行的。
小肠内转运单糖的主要系统为小肠腔上表皮细胞膜内的Na+-单糖协同转运系统,其主要功能是转运D-葡萄糊和D-半乳糖。还有一个不需Na+的易化扩散系统,主要对D-果糖有特殊的转运活性。
D-葡萄糖和D-半乳糖离开小肠表皮细胞进循环系统是通过胞浆膜中的易化扩散系统。
D-果糖则通过被动扩散作用而被转运到细胞膜。
(一)单纯扩散吸收
糖的主要吸收方式(戊糖仅靠此途径吸收)。
特点:不消耗能量、无载体参加、顺浓度梯度。
各种单糖的吸收速率不同(D-葡萄糖的吸收速率为100,如下表):
| D半乳糖110 | D甘露糖19 |
| D葡萄糖100 | L木酮糖15 |
| D果糖43 | L阿拉伯糖9 |
(二)易化扩散吸收
果糖的主要吸收方式是易化扩散吸收。
特点:不消耗能量、需要载体蛋白、顺浓度差。
速率介于单纯扩散和主动吸收二者之间。
(三)主动吸收
特点:消耗能量、需要载体蛋白、逆浓度梯度。伴Na+的顺浓度梯度同向转运。
载体蛋白对单糖的结构有选择性:
要求吡喃型单糖、并且C2上羟基同葡萄糖。(故半乳糖和葡萄糖吸收快)
Na+的浓度梯度依赖钠泵、消耗ATP。钠泵使得肠细胞内钠浓度低于肠腔钠浓度。
小肠上皮细胞膜内的Na+-单糖协同转运系统(Na+-依赖型葡萄糖转运体,SGLT)。
四、血液中的糖进入不同的组织细胞:
依赖葡萄糖转运体(glucose transportor ,GLUT)。
现在发现的GLUT有五类(GLUT1-5),分别在不同的组织细胞中发挥作用。
1、GLUT-1存在于红细胞;
2、GLUT-4主要存在于脂肪和肌肉组织。
葡萄糖在不同组织细胞的代谢途径有所不同,且分解代谢方式受氧气供应情况的影响。
五、糖代谢概况
糖代谢可分为糖的分解与糖的合成两方面。
1、糖的分解代谢包括
酵解――糖的共同分解途径;
糖有氧氧化――糖分解的主要途径;
三羧酸循环――糖的最后氧化的途径。
2、糖的合成途径包括
糖异生――非糖物质形成糖的途径;
糖原合成;
结构多糖的合成。
3、糖的其他代谢
磷酸戊糖途径(分解代谢的旁路),糖醛酸途径等。
糖代谢受神经、激素及别构物的调节控制。代谢紊乱会引起各种疾病。
第二节 糖的无氧分解
人类各种生命活动能量的来源主要是靠糖的分解代谢而提供。
糖酵解作用就是生物界的一种最古老最普通的供能方式。
所谓糖酵解作用:
1、广义地说:
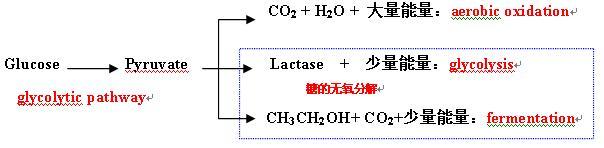
是指葡萄糖转变成丙酮酸并伴有ATP生成的序列反应过程。存在于人类及动植物界和许多微生物体内。
在需氧生物界,此酵解作用是三羧酸循环和电子传递链的前奏,从它们可以获得远比酵解作用多得多的能量。
在有氧条件下,丙酮酸进入线粒体而被完全氧化成二氧化碳和水,同时放出大量能量。但当氧供应不足时,丙酮酸则被还原成乳酸,仅能释放少许能量。
在无氧条件下,酵母等厌氧生物体既不能使丙酮酸还原成乳酸,也不能使之氧化成二氧化碳及水,而是转变成乙醇,同时也只能产生少许能量。
2、狭义地说:
葡萄糖及糖原经系列反应转变成乳酸的过程称为糖酵解作用。
葡萄糖酵解成乳酸时放出的能量,仅相当于其氧化成二氧化碳和水时放出能量的6.9%,是一种很不经济的供能方式。但是,当机体处于缺 氧的特殊生理状态时,其能量来源却主要靠糖的酵解作用;加之,糖酵解途径又是糖有氧氧化过程的必经之路。因此,酵解作用在整个糖代谢中占有相当重要的地位。
糖酵解(glycolysis):
在缺氧的情况下葡萄糖生成乳酸的过程。
糖酵解的代谢反应过程分两阶段:
1、酵解途径(glycolytic pathway):葡萄糖分解成丙酮酸的过程。
2、乳酸(lactase)的生成:丙酮酸的还原。
糖酵解的全部过程都在胞浆进行。整个过程中虽有氧化反应,但不需要氧的参加
发酵作用与酵解作用的区别:
1、发酵作用(fermentation) ―葡萄糖转变成乙醇。
为酵母及其他厌氧微生物体中的一种糖代谢过程;
发酵过程的起始物质为葡萄糖,终产物为乙醇及CO2。
2、酵解作用
人类、动植物及某些微生物体内糖代谢的一种过程;
酵解过程的起始物质为糖原或葡萄糖,终产物为乳酸。
糖的无氧分解:
发酵与酵解两种不需要氧的糖代谢过程的统称。
两个过程的起始及最终产物不同,但经历的中间化学反应步骤绝大部分完全相同,且都产生少量的能量。
有氧氧化及酵解与发酵作用如下图:
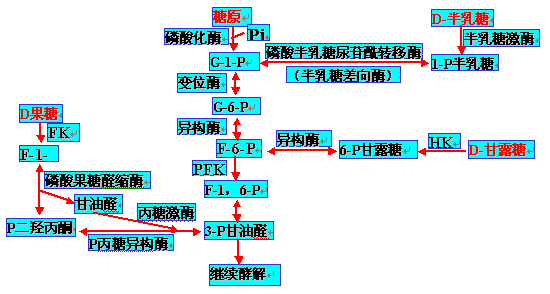
一、糖酵解的反应过程
第一阶段――葡萄糖分解成丙酮酸
(1)一分子含6碳原子的葡萄糖,经磷酸化后转变成两分子各含3个碳原子的3-磷酸甘油醛;这一阶段有两个起爆反应,是耗能过程,消耗2分子ATP,共包含5步反应。
(2)两分子3-磷酸甘油醛转变成两分子丙酮酸。这一阶段是产能过程,生成4分子ATP,共包括5步反应。
两个阶段的总结果是产能过程,净剩2分子ATP。
第二阶段――丙酮酸被NADH还原生成乳酸。
(一)第一阶段――葡萄糖生成丙酮酸,共计10步反应:有部分能量生成。
为糖无氧分解与有氧氧化的共同途径,称为酵解途径(glycolytic pathway);
区别于糖酵解(glycolysis)。
1、葡萄糖磷酸化为6-磷酸葡萄糖(glucose-6-phosphate,G-6-P):
葡萄糖进入细胞后的第一步反应;磷酸化后的葡萄糖即不能自由透过细胞膜而逸出细胞。
(1)反应过程如下:葡萄糖磷酸化为6-磷酸葡萄糖(α-D-吡喃葡萄糖)
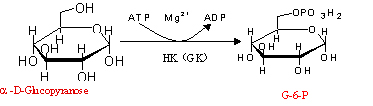
(2)反应过程的酶:己糖激酶(hexokinase,HK)
凡是催化磷酰基键ATP分子转移到受体上的酶都称为激酶。
己糖激酶的分子量为52 000,以6碳糖为底物,专一性差:
可以作用于D-葡萄糖、D-果糖和D-甘露糖。
己糖激酶像其他激酶一样,需要Mg2+或其他二价金属如Mn2+的活化。
实际上Mg2+与ATP形成的复合物才是酶的真正底物。
此酶催化的反应是糖酵解的首个调节步骤。
ATP与Mg2+的结合模式:

己糖激酶及其同工酶:
(1)肌肉己糖激酶是别构酶,被G-6-P强烈抑制。
(2)肝细胞中的HK的同工酶是Ⅳ,即葡萄糖激酶(glucokinase,GK)。
它对D-葡萄糖有特异活性,不被G-6-P抑制。
GK是诱导酶,可以被胰岛素诱导合成。GK受激素的调控是其特点。
GK对葡萄糖的亲和力较低,Km=10mmol/L(其它HK在0.1mmol/L左右)。
GK的特性决定其在维持血糖水平以及在糖代谢的调控中起重要的作用。
催化的反应不可逆,耗能反应(ΔGO/=-16.8kJ/mol,-4.0kcal/mol)。
2、6-磷酸葡萄糖转为6-磷酸果糖(fuctose-6-phosphate,F-6-P)
在磷酸己糖(葡萄糖)异构酶的催化下,G-6-P转变为F-6-P(醛酮异构)。
由己醛糖变成其异构体己酮糖的反应,分子中的羰基由第1位碳原子移位至第2位碳原子。
此反应之标准自由能变化甚小(ΔGO/=+1.68kJ/mol,+0.4kcal/mol),故反应随着作用物及产物浓度的变化,可以向正反两个方向进行。
吡喃葡萄糖与呋喃果糖
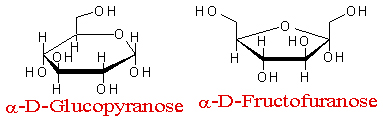
己醛糖与己酮糖的互变

3、F-6-P转变为F-1,6-2P(FBP)
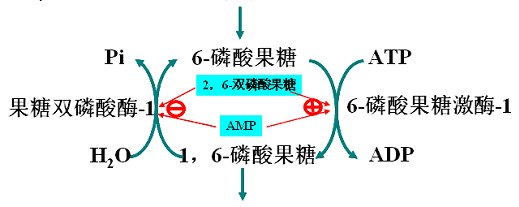
酵解过程的第2个磷酸化反应。反应由6-磷酸果糖激酶-1(PFK-1)催化。
反应是非平衡反应,倾向于生成FBP。此步反应是酵解的第二个调节反应。
F-6-P转变为F-1,6-2P(FBP)。所需要能量及其磷酸基由ATP提供。
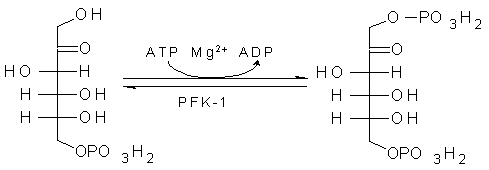
磷酸果糖激酶(PFK):
(1)分子量为3400的四聚体的别构酶。
(2)ATP对此酶有抑制效应。在有柠檬酸、脂肪酸时加强抑制效应。
AMP、ADP或Pi可消除抑制。
(3)产物1,6-FBP和2,6-FBP是此酶的激活剂。及其少见的正反馈。
此酶活力决定酵解的速度,因此是酵解中的最关键的酶,也是限速酶。
4、磷酸己糖裂解为2个磷酸丙糖
一个醛和一个酮化合物形成一个醛醇化合物就是醛缩反应,反应可逆,方向有利于磷酸己糖的合成。故催化此反应的酶称为――醛缩酶。
但是正常生理条件下,由于3-磷酸甘油醛不断转化成丙酮酸,大大降低了细胞中的3-磷酸甘油醛的浓度,从而驱动反应向裂解方向进行。
醛缩酶的反应机理:
是酶的活性中心上特殊的赖氨酸残基上的ε-氨基与磷酸二羟丙酮上的羰基形成一个四面体中间物,然后去水变成质子化的Schiff氏碱又称酮亚胺,它促使磷酸二羟丙酮形成烯醇化的负碳原子,与3-磷酸甘油醛形成质子化的Schiff碱(F-1,6-2P酮亚胺),然后再去质子化和水解陈去Schiff氏碱产生F-1,6-2P和酶。1,6-二磷酸果糖的裂解就是这个反应的逆转。
动物组织中的醛缩酶有多种同功酶,肌肉中主要是A型,肝中主要是B型,脑中是O型的,它们对金属离子或其他辅助因子无要求,而来自酵母和许多种细菌的酶则被Fe2+、Co2+或Zn2+激活,被这些金属离子的结合剂抑制。
FBP裂解为磷酸二羟丙酮和3-磷酸甘油醛――磷酸己糖裂解为2个磷酸丙糖
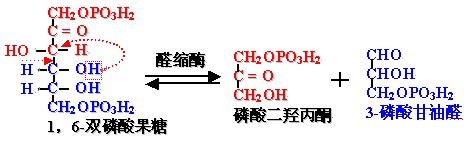
已糖转化成三碳糖后,碳原子顺序数颠倒,已糖原来的碳原子:
C3、C4→C1;
C5、C2→C2;
C1、C6→C3;
5、磷酸丙糖的互变
两个磷酸丙糖(磷酸二羟丙酮和3-磷酸甘油醛)是可以互变的同分异构体。
磷酸丙糖的互变由磷酸丙糖异构酶催化,方向利于磷酸二羟丙酮的生成;平衡时占96%。
由于3-磷酸甘油醛不断继续酵解被移去,故最终磷酸二羟丙酮全部转变为3-磷酸甘油醛。

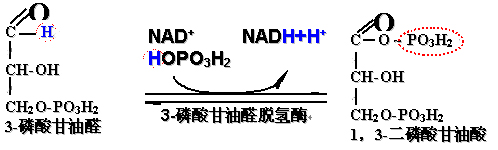
6、3-磷酸甘油醛氧化成1,3-二磷酸甘油酸
(1)此反应包括:
醛基的氧化及羧基磷酸化两步反应,均由3-磷酸甘油醛脱氢酶催化;
反应需要NAD+为辅酶受氢及电子;
磷酸化反应需要无机磷酸参与。
(2)反应生成的1,3-二磷酸甘油酸为含有高能磷酸键的混合酸酐:
1,3-二磷酸甘油酸水解时自由能变化为:
ΔG0/= -14.8千卡/摩尔(-61.9千焦耳/摩尔);可以用来进行ADP的磷酸化生成ATP。
这个反应既是氧化反应又是磷酸化反应。如需要将磷酸甘油醛上C1醛基上的氢质子和电子移去,则有很大的能量障碍,因为羰基的碳原子已经带正电,为此必须加入另一亲核试剂,以减少羰基碳上的正电性,使羟基上的氢能够移去,则磷酸加入后容易形成酰基磷酸,后者是羰基与磷酸的混合酸酐,具有强烈的转移磷酰基的能量。
磷酸甘油醛脱氢酶活性位置上半胱氨酸残基的巯基就是亲核基团,它与醛基作用形成中间化合物可将羟基上的氢移至与酶紧密结合的NAD+上,从而产生NADH和高能硫酯中间物。然后NADH从酶上解离下来,另外的NAD+与酶活性位置结合,磷酸攻击硫酯键就形成1,3-DPG。
酯酰磷酸化反应是热力学不利的反应,其ΔG0′ =49.56千焦耳/摩尔(+11.8千卡/摩尔),但是氧化反应的ΔG0′ =-43.2千焦耳/摩尔(-10.3千卡/摩尔)。两个反应由硫酯中间物而偶联在一起,氧化反应的能量驱动磷酸化反应进行。
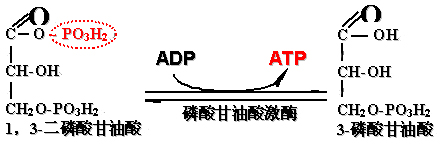
7、1,3-二磷酸甘油酸转变成3-磷酸甘油酸
底物水平磷酸化:
ADP或其它核苷二磷酸的磷酸化作用与底物的脱氢作用直接偶联的反应过程。
此步反应是糖酵解过程的第一个产生ATP的反应:
磷酸甘油酸激酶催化混合酸酐的磷酸从羧基转移到ADP生成ATP和3-磷酸甘油酸。
1,3-二磷酸甘油酸转变成3-磷酸甘油酸伴ADP磷酸化
8、3-磷酸甘油酸转变成2-磷酸甘油酸――同分异构体之间的转变:
反应可逆,并且需要Mg2+参加。磷酸甘油酸变位酶催化3-磷酸甘油酸的磷酸基从C3转移到C2。

反应机理
凡是促化分子内化学功能基团的位置移动的酶都称为变位酶。Mg2+在催化反应中是必需的。
ΔG0′=+1.06千卡/摩尔=+4.45千焦耳/摩尔。
反应需要2,3-DPG为辅助因子,其反应机理是:
E-P + 3-PG ←→ E + 2,3-DPG ←→ E-P + 2-PG
哺乳动物中至少有二种磷酸甘油酸变位酶。
酶的分子量是65700,其一存在于成人肌肉,另存在于胎儿肌肉。前者对于汞高度敏感。
9、2-磷酸甘油酸转变成磷酸烯醇式丙酮酸
烯醇化酶催化2-磷酸甘油酸脱水成PEP。
此反应引起分子内电子重排和能量重分布,形成一个高能磷酸键,为下一步底物水平磷酸化作准备。

10、PEP转变成丙酮酸;伴有ADP的磷酸化生成ATP
糖酵解反应的第二次底物水平磷酸化。反应由PK催化;需要K+、Mg2+参加;细胞内此反应不可逆。

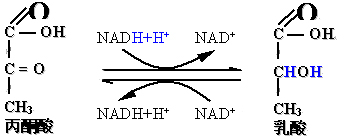
(二)第二阶段――丙酮酸还原成乳酸:
乳酸脱氢酶(LDH)催化的可逆反应:反应方向由供氧情况决定。辅酶是NAD+(NADH+H+)。
葡萄糖酵解途径的总反应式:
葡萄糖+2Pi+2ADP+2NAD+→2丙酮酸+2ATP+2NADH+2H++2H2O
无氧情况下糖酵解的反应式:葡萄糖十2Pi十2ADP→2乳酸十2ATP十2H2O
糖酵解全过程:

糖酵解共产生2分子ATP,2分子NADH将H交给2分子丙酮酸生成2分子乳酸。
注意:有氧情况下2分子NADH经呼吸链氧化成H20可产生6(4)分子ATP,故1分子葡萄糖酵解过程共产生8(6)分子ATP。
不同生物的不同条件下,丙酮酸有不同的去路
1、变为乙酰CoA(有氧条件)
丙酮酸进入线粒体变成乙酰CoA参加三羧酸循环,最后氧化成CO2和H2O。
2、生成乳酸(缺氧条件)
在厌氧酵解、肌肉剧烈运动、呼吸循环系统机能障碍暂时造成暂时缺氧时,丙酮酸接受3-磷酸甘油醛脱下的氢,还原成乳酸。
3、生成乙醇(酵母菌或其他微生物)
丙酮酸经脱羧酶催化(TPP辅酶)脱羧成乙醛后醇脱氢酶催化下由NADH还原形成乙醇。
二、糖酵解的调节
糖酵解途径多数反应可逆:
(1)可逆反应方向和速率由底物和产物浓度控制。
(2)可逆反应酶活性的变化不能决定反应的方向。
糖酵解途径有三个酶催化三个不可逆反应,是糖酵解途径流量的三个调节点,受变构剂和激素的调节。
(1)己糖激酶(葡萄糖激酶)――HK(GK)
(2)6-磷酸果糖激酶-1(PFK)
(3)丙酮酸激酶(PK)
(一)6-磷酸果糖激酶-1:调节糖酵解途径流量最重要的酶。
1、多种变构效应剂:
(1)变构抑制剂:ATP、柠檬酸
酶分子有2个ATP的结合部位(催化部位和别位)
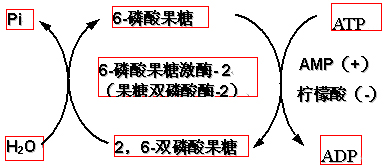
(2)变构激活剂:Pi、AMP、ADP、F-2,6-2P、F-1,6-2P
AMP与ATP竞争结合部位。F-2,6-2P、F-1,6-2P属于产物正反馈。
F-2,6-2P是6-磷酸果糖激酶-1最强的激活剂。
在生理浓度(微摩尔水平)即可发挥作用。
2,6-双磷酸果糖的生成与脱磷酸都由6-磷酸果糖激酶-2(果糖双磷酸酶)催化。
6-磷酸果糖激酶-2与果糖双磷酸酶实际上是一种酶同时具有两种活性,属于双功能酶。
(3)此酶可以在激素作用下通过共价修饰调节酶活性。
如:胰高血糖素通过cAMP和PKA使其磷酸化。磷酸化的结果是――激酶活性减弱、磷酸酶活性提高。2,6-双磷酸果糖的合成与分解如下:
(二)丙酮酸激酶
糖酵解途径的第二重要的调节点。
1、变构调节:
(1)F-1,6-BP和PEP是变构激活剂;
(2)ATP(肝内丙氨酸)、柠檬酸、乙酰CoA是变构抑制剂。
2、共价修饰调节:
依赖cAMP、Ca2+、CaM的蛋白激酶使其磷酸化失活。
如:胰高血糖素通过cAMP抑制丙酮酸激酶。
PK活性的调控
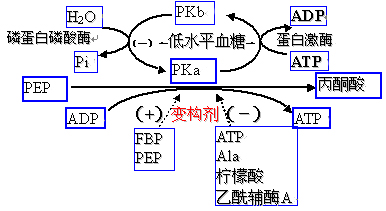
(三)己糖激酶(葡萄糖激酶)
作用不及前二者重要。
1、变构抑制剂:
(1)HK受反应产物G-6-P的反馈抑制(GK不受G-6-P的反馈抑制);
(2)长链脂酰CoA是变构抑制剂;
2、胰岛素的作用:
胰岛素诱导GK基因的转录、促进酶合成。
三、糖酵解的生理意义
1、最主要意义――迅速供能,对肌肉收缩最重要。
比有氧氧化来的迅速(即使氧气供应充足,但时间较长)。
肌肉运动或机体缺氧时肌肉主要靠酵解供能。
(1)1mol葡萄糖酵解净剩2molATP,可以储存能量为30.5×2=61kJ/mol(14.6kcal/mol);(以标准状态ATP水解的ΔGO/=-30.5kJ/mol,-7.29kcal/mol计):
所以储存能量的效率=14.6/47=31%。
(2)生理条件ATP水解的自由能变化高于标准状态,ΔG=-51.6kJ/mol(-12.3kcal/mol),所以储存能量的效率为24.7/47=52.6%。
注:
此阶段(Glucose+2Pi+2ADP→2Lactate+2ATP+2H2O)自由能变化:
ΔGO/=-32.4kcal/mol(释放的能量以热能形式散发);
释放的热能+ATP储存的化学能=32.4kcal/mol+14.6kcal=47kcal
1molGlucose完全氧化为水和二氧化碳释放能量2840kJ(679kcal),经糖酵解仅释放少量自由能,用于合成ATP及以热能散发,大多数能量储存在乳酸中(679-47=632kcal,占葡萄糖蕴藏总能量的93%)。
2、特殊细胞只能利用糖酵解供应能量。
(1)红细胞:
无线粒体,完全靠糖酵解供能。
(2)其它组织:
如神经、骨髓、白细胞代谢活跃,即使不缺氧也常由糖酵解提供部分能量。
3、糖酵解在体内物质互变过程中的重要作用:
丙酮酸可转变成丙氨酸,丙氨酸可以转变成其它一些氨基酸;
糖酵解过程中生成的磷酸丙糖可转变成α磷酸甘油和甘油,是体内合成脂肪的原料。
蛋白质、脂肪等可转变为糖酵解途径的中间产物,在肝中转变为糖原和葡萄糖。
各种己糖进入糖酵解的途径
第三节 糖的有氧氧化
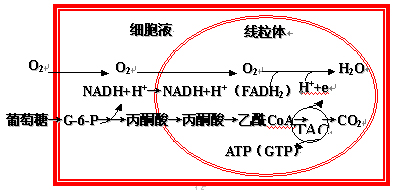
有氧氧化(aerobic oxidation):
葡萄糖在有氧条件下彻底氧化成水和二氧化碳的反应过程。
有氧氧化是糖氧化的最主要方式,是绝大多数细胞利用葡萄糖获取能量的方式。
肌肉等组织酵解生成的乳酸最终仍需要在有氧的条件下彻底氧化成水和二氧化碳。
葡萄糖有氧氧化概况
一、有氧氧化的反应过程
糖有氧氧化分三个阶段:
第一阶段――葡萄糖经糖酵解分解成丙酮酸(见糖酵解)
第二阶段――丙酮酸进入线粒体氧化脱羧成乙酰CoA
第三阶段――三羧酸循环及氧化磷酸化(见生物氧化)
(一)丙酮酸氧化脱羧
葡萄糖酵解生成的丙酮酸进入线粒体氧化脱羧生成乙酰CoA:
1、总反应式:
丙酮酸+NAD++HSCoA→乙酰CoA+NADH(H+)+CO2
2、催化反应的酶:
丙酮酸脱氢酶系(复合体)――三种酶、五种辅助因子
(1)丙酮酸脱氢酶(E1)――TPP
(2)二氢硫辛酰胺转乙酰酶(E2)――硫辛酸、CoA
(3)二氢硫辛酰胺脱氢酶(E3)――FAD、NAD+
真核细胞(哺乳类动物)的丙酮酸脱氢酶复合体存在于线粒体中;
由三种酶按不同比例(随生物体而异)组成:
(1)60个转乙酰酶(E2)组成核心;
(2)12个丙酮酸脱氢酶(E1);
(3)6个二氢硫辛酰胺脱氢酶(E3)。
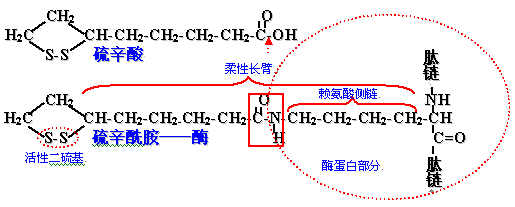
硫辛酸:
带有二硫键的八碳羧酸,通过羧基与转乙酰酶的赖氨酸ε-氨基结合成硫辛酰胺-酶;
其硫辛酰胺成为酶的柔性长臂,可以将乙酰基从酶复合体的一个活性部位转移到另外一个活性部位。硫辛酸及其硫辛酰胺的结构如下:
3、反应过程
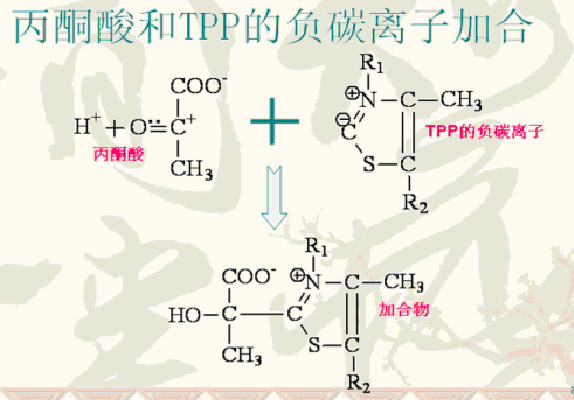
(1)丙酮酸脱羧成羟乙基-TPP
反应式:即丙酮酸十TPP→羟乙基-TPP十CO2
催化此反应的酶:是“丙酮酸脱氢酶组分”(E1),其辅酶为TPP。
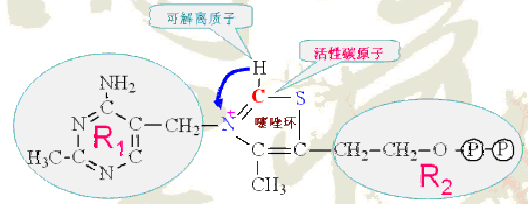
TPP的特点:
其分子中噻唑环上氮原子和硫原子之间的碳原子具酸性,可以解离成负碳离子;
此负碳离子很易加到丙酮酸的碳基上形成中间加合物,其带正电荷的含氮环作为电子阱稳定负电荷的形成,这种作用是脱羧反应所必需的;
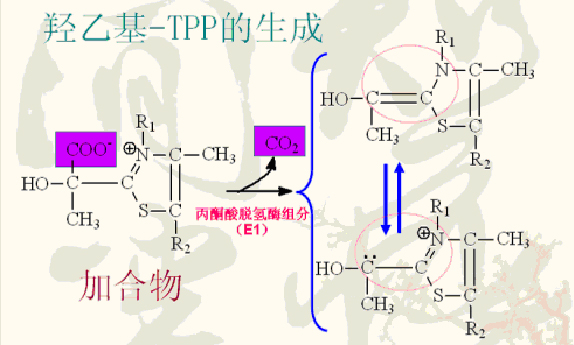
加合物脱羧后则生成羟乙基-TPP。
TPP的结构
丙酮酸和TPP的负碳离子形成中间体加合物。
(2)羟乙基氧化及其转移
羟乙基-TPP-E1中的羟乙基被氧化成乙酰基并立即转移给硫辛酰胺生成乙酰硫辛酰胺。
催化此反应的酶也是复合体中的丙酮酸脱氢酶组分(E1)。
此反应的氧化剂为硫辛酰胺中的二硫基。
反应中放出的能量集中于与乙酰基相连的高能硫酯键上。
二硫基被还原成硫氢基(巯基)形式。
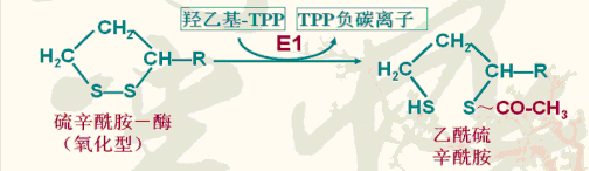
(3)乙酰辅酶A生成
乙酰硫辛酰胺中的乙酰基连同高能硫酯键转移给辅酶A,生成乙酰CoA。
催化此反应的酶:二氢硫辛酰转乙酰基酶(E2)

(4)二氢硫辛酰胺氧化脱氢
还原型二氢硫辛酰胺在二氢硫辛酰胺脱氢酶(E3)催化下脱氢:FAD接受氢还原成FAD?2H。FAD?2H将氢原子再转移给NAD+,生成还原型NADH+H+。
氧化型的硫辛酰胺得以再生被重新利用。
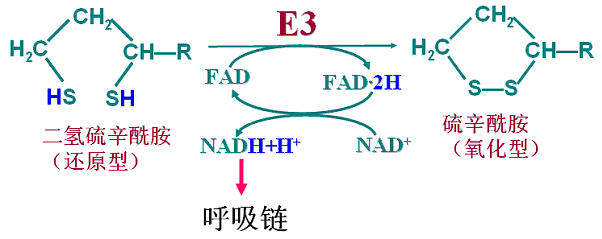
丙酮酸氧化脱羧总过程

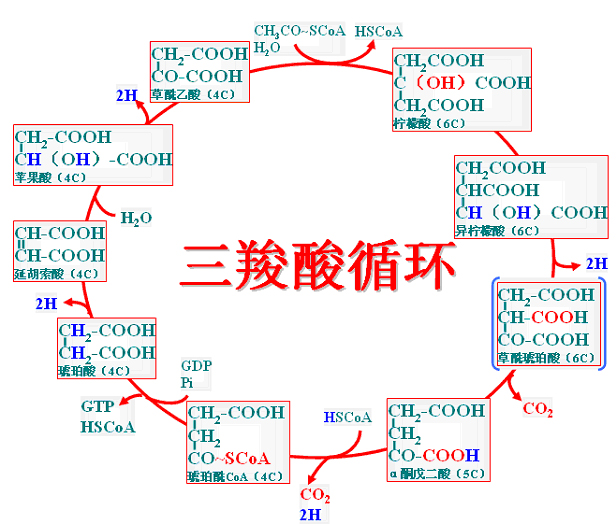
(二)三羧酸循环
三羧酸循环(tricarboxylic acid cycle)、柠檬酸循环(citrate cycle)或Krebs循环(Krebs cycle),多种酶催化的连续的化学反应。
1、三羧酸循环的反应过程
(1)柠檬酸的形成
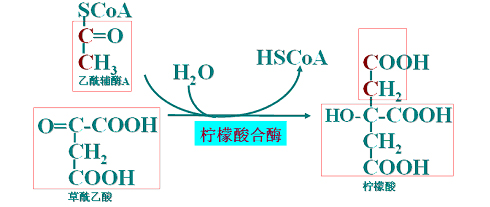
(2)异柠檬酸生成
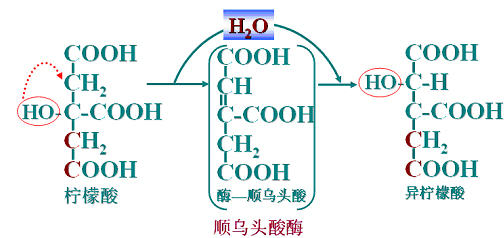
(3)第一次氧化脱羧
异柠檬酸脱氢酶(NAD+为辅酶)催化,此反应不可逆。(先脱氢成草酰琥珀酸再脱羧)

(4)第二次氧化脱羧
α-酮戊二酸脱氢酶复合体(类似PDH)催化α-酮戊二酸脱氢、脱羧、形成高能硫酯键。
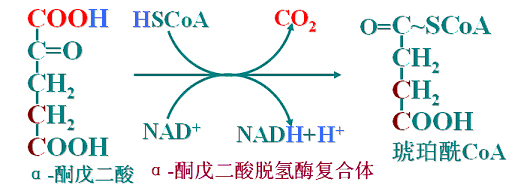
(5)底物水平磷酸化
琥珀酰CoA合成酶催化的可逆反应,TAC中唯一达到一次底物水平磷酸化。

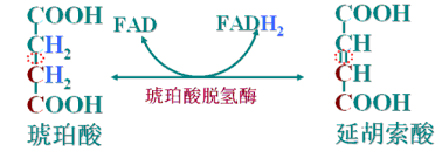
(6)琥珀酸脱氢成延胡索酸
琥珀酸脱氢酶(FAD为辅酶)是TAC中唯一与线粒体内膜结合的酶。
琥珀酸脱下的氢可以直接进入电子链氧化。
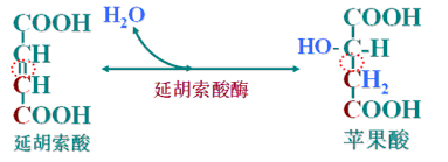
(7)延胡索酸加水成苹果酸
延胡索酸酶催化的加水与脱水的可逆反应。
(8)苹果酸脱氢生成草酰乙酸
苹果酸脱氢酶催化的可逆反应。由于草酰乙酸不断用于合成柠檬酸;所以反应向生成草酰乙酸的方向进行。

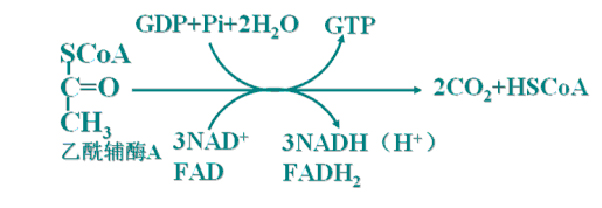
2、三羧酸循环的总反应
加水缩合为柠檬酸开始:4次脱氢(2次伴有脱羧)――3NAD+、1FAD、1次底物水平磷酸化(GTP)。
总反应式:
说明
(1)TAC中表面上一个二碳化合物被氧化成2分子CO2,但是CO2中的碳实际来自草酰乙酸而不是乙酰CoA;
原因在于顺乌头酸酶的构象对于柠檬酸的构象有选择性。
柠檬酸虽然是对称分子,但是以中间碳原子为中心形成一个正四面体,并且与顺乌头酸有三个不同作用位点(从中心碳顺时针观察):
羟基-羰基-未标记乙酰基(唯一正确的作用位点与酶吻合):![]()
羟基-未标记乙酰基-标记乙酰基:![]()
羟基―标记乙酰基-羰基:![]()
在主体位置上,乙酰CoA的乙酰基和草酰乙酸的乙酰基不一致,是由于顺乌头酸酶与柠檬酸的结合不对称,脱水时氢原子只能来自草酰乙酸部分。

(2)TAC中的中间产物包括草酰乙酸起催化剂的作用,本身无量的消耗,但是其中的C被乙酰CoA置换;
TAC的中间物不可能直接被氧化;也不是被合成。
3、三羧酸循环的生理意义
(1)三羧酸循环是三大营养素的最终代谢通路:
糖、脂肪、氨基酸的氧化都产生乙酰辅酶A,然后进入TAC降解。
1次底物水平磷酸化产生高能磷酸键;
更加重要的是通过4次脱氢为氧化磷酸化提供还原当量。
(2)三羧酸循环是三大营养素代谢联系的枢纽
糖→丙酮酸→乙酰CoA→脂肪酸→脂肪;乙酰CoA→胆固醇
氨基酸→TAC中间物→草酰乙酸→糖
葡萄糖→丙酮酸→草酰乙酸等二羧酸→非必需氨基酸
(3)为其他生物合成提供前体
琥珀酰CoA→血红素。
二、糖有氧氧化的意义
1.供能并提供合成其它物质的原料
2.通过TCAC同其它物质代谢相联系并产生二氧化碳和内源性水。
3.与糖代谢的其它途径联系的枢纽,是糖代谢的干线。
三、有氧氧化的能量代谢――(ATP)的生成
反应 |
辅酶 |
ATP |
| 葡萄糖→G-6-P | -1 | |
| F-6-P→F-1,6-P | -1 | |
| 3-P甘油醛→1,3-DPG×2 | NAD+ | 3*或2* ×2 |
| 1,3-DPG→3-P甘油酸×2 | 1 ×2 | |
| PEP→丙酮酸×2 | 1 ×2 | |
| 丙酮酸→乙酰辅酶A×2 | NAD+ | 3 ×2 |
| 异柠檬酸→α-酮戊二酸×2 | NAD+ | 3 ×2 |
| α-酮戊二酸→琥珀酰辅酶A×2 | NAD+ | 3 ×2 |
| 琥珀酰辅酶A →琥珀酸 ×2 | 1×2 | |
| 琥珀酸 →延胡索酸×2 | FAD | 2×2 |
| 苹果酸→草酰乙酸×2 | NAD+ | 3 ×2 |
| G+38ADP+38Pi+6O2→38ATP+6CO2+44H2O | 38或36 |
四、有氧氧化的调节
(一)糖酵解的三个关键酶(见前述)
(二)丙酮酸脱氢酶复合体(PDH)的调节
1、反馈调节
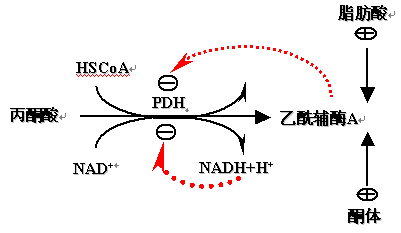
2、酶的共价修饰
活性的PDH-a型及无活性的PDH-b型两种形式:

PDH激酶可受多种因素的调节:
(1)Ca2+、二氯乙酸、丙酮酸等可抑制此酶的活性,
(2)ATP/ADP、NADH/NAD+、乙酰CoA/CoA增加可激动此酶的活性。
PDH磷酸酶使PDH-b脱磷酸而变成PDH-a。
(1)在脂肪组织中可被胰岛素激活
(2)Mg2+及Ca2+亦有同样的激活作用。
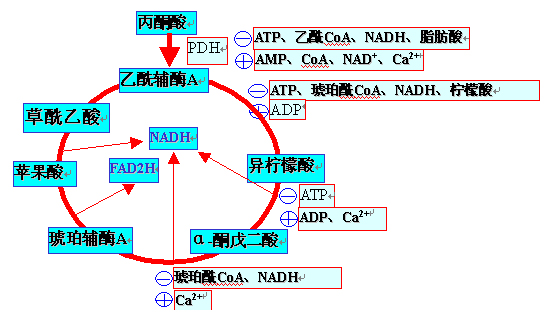
(三)TAC的调节:
三个不可逆反应(柠檬酸合成、异柠檬酸脱氢、酮戊二酸脱氢),后两个是主要调节点。
还存在代谢途径间的作用(巴斯德效应及反效应)
1、DNAH/NAD+、ATP/ADP比率增高:
反馈抑制异柠檬酸脱氢酶和α-酮戊二酸脱氢酶;
ADP是异柠檬酸脱氢酶的变构激活剂。
2、Ca2+浓度升高:
可以降低其对底物的Km
激活异柠檬酸脱氢酶和α-酮戊二酸脱氢酶。
3、氧化磷酸化的速率也影响TAC的运转。
三羧酸循环的调控
(四)巴斯德(Pastuer)效应
1、巴斯德效应:有氧氧化抑制糖酵解的现象。
有氧时NADH+H+可以进入线粒体氧化,丙酮酸也不转变为乳酸,而进一步氧化、进入三羧酸循环。
2、反巴斯德效应:糖无氧酵解抑制糖有氧氧化的现象。
缺氧时NADH+H+不能进入线粒体氧化,丙酮酸成为受氢体转变为乳酸。
第四节 磷酸戊糖途径(pentose phosphate psthway)
糖分解的途径,又称磷酸戊糖旁路或磷酸己糖旁路(hexose monophosphte shunt,HMP)、Warburg―Dikens 途径。
部位:胞浆。注:此途径尽管是糖的分解途径,但主要意义不是产生ATP。

一、磷酸戊糖途径的反应过程
反应过程(2阶段),整个反应不可逆:
氧化反应:产生磷酸戊糖、二氧化碳和还原性辅酶II
基团转移反应:产生不同碳原子数的磷酸单糖。
(一)磷酸戊糖的生成
1、氧化阶段――生成三种产物:
(1)1分子磷酸戊糖(可以有3种异构体);
(2)2分子NADPH+H+;
(3)1分子CO2。
生成的三种磷酸戊糖互为异构体:
(1)5-磷酸核酮糖
(2)5-磷酸木酮糖(5-P核酮糖的差向异构体)
(3)5-磷酸核糖(5-P核酮糖的醛酮异构体)
(二)基团转移反应――产生不同碳原子数的单糖
最终核糖可以转变为:2分子6-磷酸果糖、1分子3-磷酸甘油醛。二者都进入糖酵解途径。
中间有4-磷酸赤藓糖和7-磷酸景天糖生成。
催化基团转移的酶:转酮醇酶和转醛醇酶,受体都是醛糖。
(1)转酮醇酶:
转移含1个酮基、1个醇基的二碳基团(羟乙醛)。
(2)转醛醇酶:
转移三碳单位(二羟丙酮)。
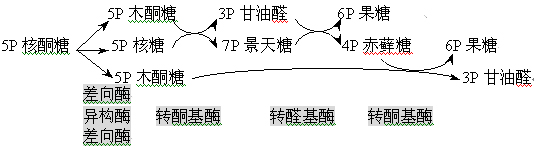
磷酸戊糖途径的总反应:
3 G6P + 6 NADP+ ----→2 F6P + 3-磷酸甘油醛 + 6NADPH(H+)+ 3 CO2
机体对于NADPH(H+)的需要大于对核糖的需要,多余的核糖转变为 F6P 和3-磷酸甘油醛进入糖酵解途径。
磷酸戊糖途径的总反应过程:

二、磷酸戊糖途径的调节
1、限速酶:6-磷酸葡萄糖脱氢酶。此酶决定G-6-P进入该途径的流量。
2、6-磷酸葡萄糖脱氢酶活性的快速调节――NADPH/NADP+比值是主要因素:
(1)NADPH/NADP+比值↑抑制此途径
(2)NADPH/NADP+比值↓激活此途径
NADPH是此酶的强抑制剂。磷酸戊糖途径的流量取决于对NADPH的需求
三、磷酸戊糖途径的生理意义
产生磷酸核糖和产生NADPH+H+
1、核糖5-磷酸是合成核苷酸和核酸及其核苷酸辅酶的原料
2、NADPH+H+作为供氢体参与体内的多种代谢反应:
(1)各种物质合成(脂肪酸、胆固醇、氨基酸等)
(2)参与体内羟化反应(物质合成、生物转化)
(3)作抗氧化剂维持谷胱甘肽的还原状态。
NADPH(H+)是谷光甘肽还原酶的辅酶,维持还原型谷光甘肽(G-SH)的含量。
(3)NADPH(H+)参与体内噬中性粒细胞和巨噬细胞产生离子态氧的反应,有杀菌作用。
(4)葡萄糖可以经过此途径生成二氧化碳并与其它代谢途径相连。
第五节 糖原的合成与分解
糖原是动物体内糖的储存形式,但只占糖的少部分,糖类大部分以脂肪形式储存。
糖原作为葡萄糖储备的意义在于机体需要葡萄糖时可以迅速动员。
糖原主要储存于肌肉和肝脏:
肝糖原的作用主要是迅速补充血糖;肝糖原含量70-100克(饥饿24小时耗尽)。
肌糖原主要是供给肌肉收缩时的能量;肌糖原含量180-300克。
糖原的结构示意图:
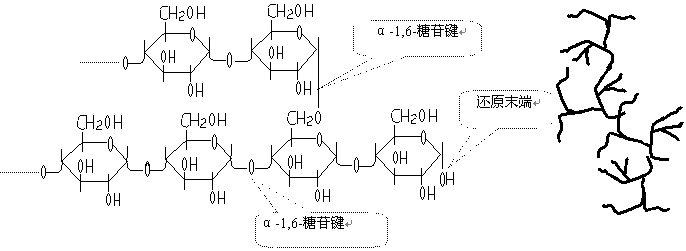
糖原的结构特点
树枝状的多聚葡萄糖。分子量100-1000万。
糖原含有一个还原性末端、多个非还原性末端(合成与分解都从此末端开始)。
直链7-12葡萄糖单位以α1,4-糖苷键(占总糖苷键的93%)连接;
直链之间以α1,6-糖苷键(7%)连接成分支。
一、糖原的合成代谢
糖原合成:
在肝脏和肌肉利用葡萄糖合成糖原。糖原合成需要糖原引物(原有的小分子),糖原合成时葡萄糖的供体是UDPG。
糖原合成包括如下几个同时进行的过程:
葡萄糖的活化(UDPG焦磷酸化酶)、直链的延长(糖原合成酶)、分支的形成(分支酶)。
(一)葡萄糖活化
此过程即形成活性葡萄糖――UDPG
1.G + ATP -----→ G6P + ADP (HK、GK)
2.G6P -----→ G1P(磷酸葡萄糖变位酶)
3.G1P + UTP -----→UDPG + PPi(UDPG-焦磷酸化酶)
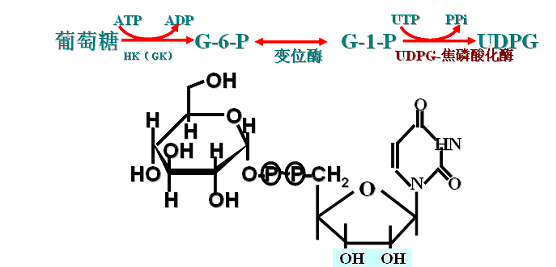
(二)糖原引物直链的延长
链的延长是从非还原性末端开始,形成α-1,4-糖苷键,每次增加一个葡萄糖单位:
糖原合成

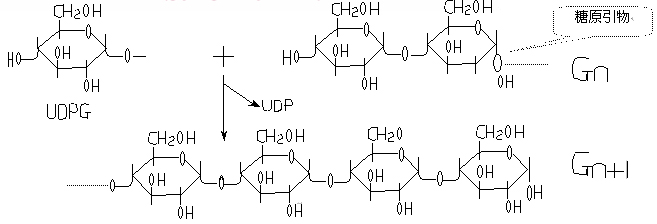
催化此反应的酶是糖原合成酶(glycogen synthetase)或UDPG-糖原葡萄糖基转移酶。
有I型和D型2种,可以互变,前者有活性。此酶是限速酶。
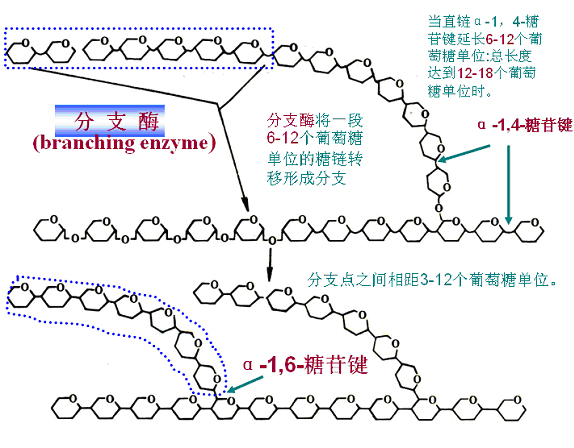
(三)分支的形成
当直链(α-1,4-糖苷键连接)延长6―12(总长度12-18)个葡萄糖单位时,分枝酶将1段含有6―(7)12个葡萄糖单位的糖链转移形成分枝(α-1,6-糖苷键连接),分枝之间相距3―12个葡萄糖单位。
说明:
糖原合成是多个糖原分子(引物)的许多非还原末端同时反复进行反应,延长糖链并形成新分枝,使糖原分子不断增大,一般不形成全新的糖原分子。
即:糖原合成是在原有引物的基础上进行的,只是大小改变,数量基本不变。

二、糖原的分解代谢
糖原分解:
糖原分解为葡萄糖或1-磷酸葡萄糖的过程。糖原分解主要是磷酸解。
糖原分解是糖原磷酸化酶和脱支酶共同作用的结果。
糖原磷酸化酶是糖原分解的关键酶。
脱支酶具有2种活性:葡聚糖转移酶和α1,6-葡萄糖苷酶。
糖原分解的过程:
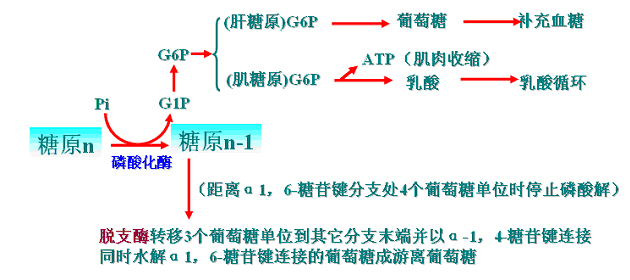
糖原分解示意图
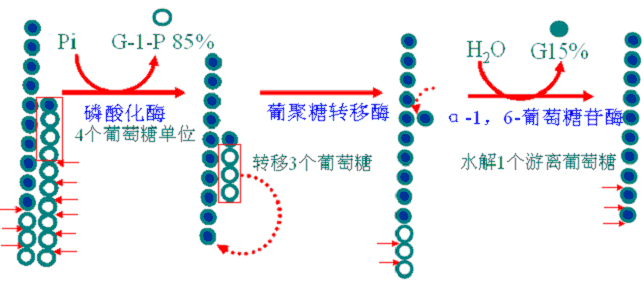
糖原合成说明:
1.糖原磷酸化酶是糖原分解的关键酶,有磷酸化酶a和磷酸化酶b两种可以互变的形式;前者有活性。产物是葡萄糖1-磷酸。
2.寡α-1,4→α-1,6-葡聚糖转移酶和α-1,6-葡萄糖苷酶是串联酶(在同一条多肽链上),故合称脱枝酶(debranching enzyme)。其作用如下。

3.肝脏中葡萄糖6-磷酸经葡萄糖6-磷酸酶作用生成葡萄糖直接补充血糖。
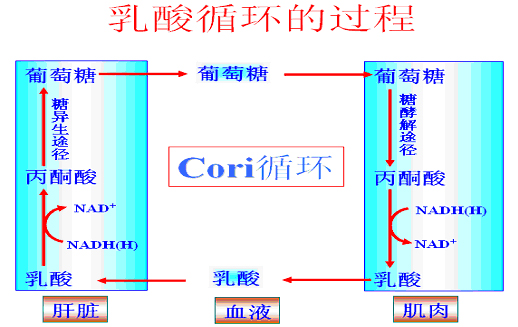
肌肉中葡萄糖6-磷酸则参加糖酵解(尤其是供氧不足时)放能供肌肉收缩,产生的乳酸则进入乳酸循环(Cori循环),间接补充血糖。(乳酸循环见糖异生)。
乳酸循环的意义在于――避免损失乳酸并防止乳酸堆积引起酸中毒。
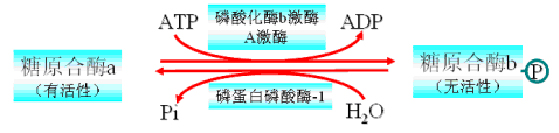
三、糖原合成与分解的调节
糖原的合成与分解是分别进行的2条途径:
糖原合成的关键酶是糖原合酶;糖原分解的关键酶是磷酸化酶
糖原合酶与磷酸化酶的活性决定糖原两条代谢途径的速度和方向。两种酶活性的快速调节方式包括:变构调节和共价修饰。
(一)磷酸化酶
1、变构调节――葡萄糖是变构抑制剂
2、共价修饰

(二)糖原合酶
共价修饰
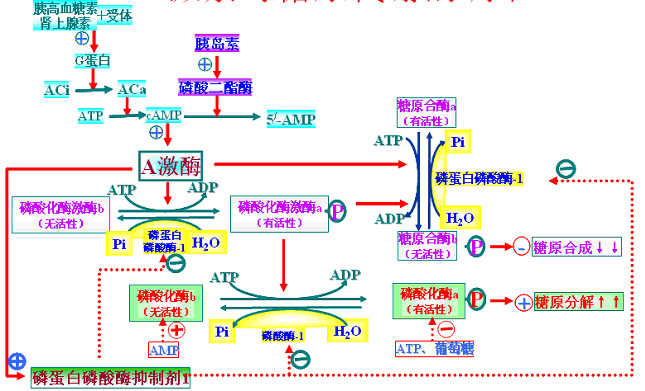
(三)激素对糖原代谢的调节
四、糖原累积症
是一种遗传性代谢病。原因是患者先天性缺乏糖原代谢相关的酶。
特点:体内某些组织器官中有大量糖原堆积。
根据缺乏或缺陷的酶的不同,受累器官和糖原结构也不同,所以对健康和生命的影响程度也不一样。糖原累积症的分型(I――X)如下表:
| 分型 | 缺陷的酶 | 受累器官 | 分型 | 缺陷的酶 | 受累器官 |
| Ⅰ | 葡萄糖6-磷酸酶 | 肝、肾 | Ⅵ | 肝糖原磷酸化酶 | 肝 |
| Ⅱ | 溶酶体α葡糖苷酶 | 所有组织 | Ⅶ | 磷酸果糖激酶-1 | 肌肉、红细胞 |
| Ⅲ | α-1,6-葡糖苷酶 | 肝、肌肉 | Ⅷ | 腺苷酸磷酸化酶? | 脑、肝 |
| Ⅳ | 分枝酶 | 所有组织 | Ⅸ | 磷酸化酶激酶 | 肌肉外组织 |
| Ⅴ | 肌糖原磷酸化酶 | 肌肉 | Ⅹ | 依赖cAMP的蛋白激酶 | 肝、肌肉 |
糖原结构:Ⅲ型分枝多,外周糖链短;Ⅳ型分枝少,外周糖链特别长。其余正常。
第六节 糖异生
肝糖原补充血糖――12小时。
肝糖原耗尽以后――糖异生补充血糖。
糖异生:
非糖化合物(乳酸、氨基酸、甘油)转变为葡萄糖或糖原的过程。
糖异生的器官:
主要是肝脏;
肾(皮质),糖异生能力是肝的1/10(注:长期饥饿时肾糖异生能力加强,几乎等于肝脏)。
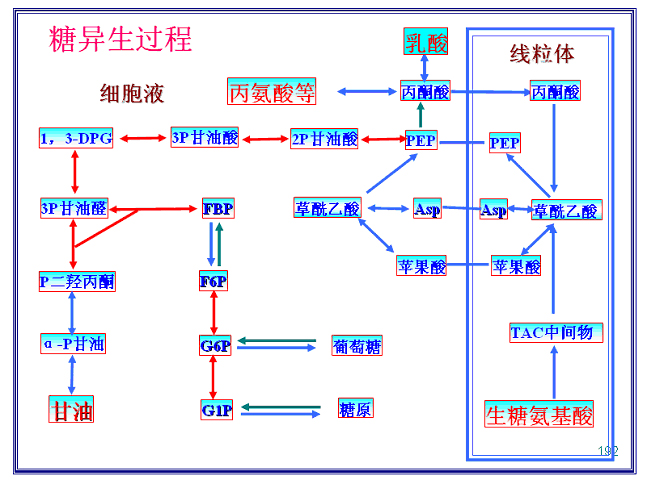
一、糖异生途径
糖异生途径:
从丙酮酸生成葡萄糖的具体反应过程;即:糖酵解的逆过程。
糖酵解与糖异生多数反应途径是共有的。
但是:
糖酵解的三个不可逆(吸能)反应由另外的反应和酶催化:底物循环和丙酮酸羧化支路。
注:丙酮酸羧化支路也可以看作是底物循环。
(一)丙酮酸→PEP
2个反应组成、由两个关键酶催化、消耗两个高能磷酸键(ATP、GTP)
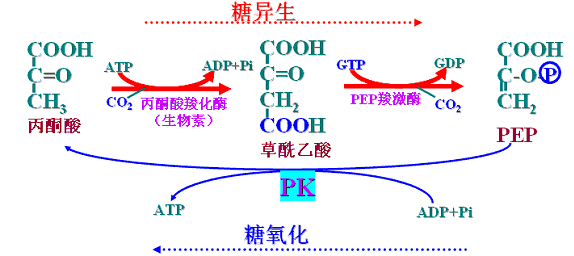
1、丙酮酸羧化酶(辅酶是生物素)――丙酮酸羧化支路
是糖异生的关键酶,仅存在于线粒体,故丙酮酸必须进入线粒体才能羧化成草酰乙酸。
反应分两步进行:
CO2与生物素结合――CO2的活化(耗能):生物素将活化的CO2转移给丙酮酸生成草酰乙酸。
2、磷酸烯醇式丙酮酸羧激酶――
糖异生的关键酶,存在于细胞液和线粒体,故草酰乙酸可以在细胞液和线粒体两处转变为PEP。反应消耗一个高能磷酸键(GTP)。反应伴有脱羧(二氧化碳)
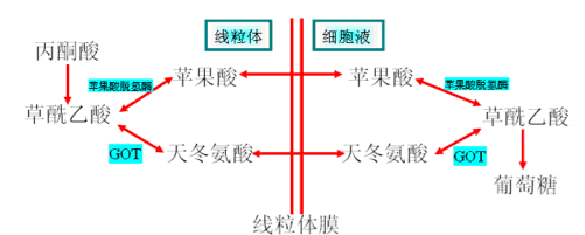
草酰乙酸出线粒体――“膜障”的克服
草酰乙酸不能直接透过线粒体膜,必须借助于两种方式:苹果酸(氨基酸)或天冬氨酸(乳酸)。
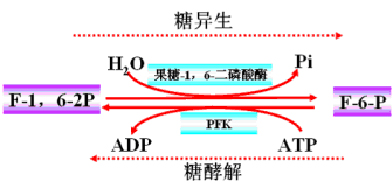
(二)F-1,6-2P→F-6-P
底物循环:PFK和果糖1,6-二磷酸酶。
C1的磷酸酯水解是放能反应,不生成ATP。反应易于进行。
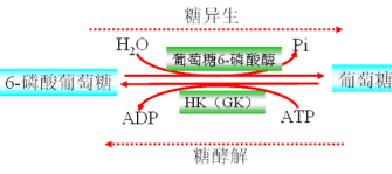
(三)G-6-P→葡萄糖
底物循环(作用物循环):HK(GK)和葡萄糖6-磷酸酶。同样不生成ATP。
底物循环(substrate cycle)
底物循环概念:
分别由不同的酶催化其单向反应,使作用物的互变的循环。
两方向酶活力相等时的循环是无效循环,不能向前推进代谢,只是ATP分解释放能量。
细胞内两向酶活力不同,代谢只能单向进行。
对两方向酶活力的调节常常相反(底物循环的两方向的酶活力变化相反)。
注意:一个单向反应可以不只一种酶。
二、糖异生的调节
糖异生的调节是对糖酵解和糖异生两个相反途径的协调调节。
若进行有效的糖异生,就必须抑制糖酵解。反之亦然。
协调调节主要针对两个底物循环:F6P←→FBP和PEP←→丙酮酸。
(一)对F-6-P――FBP的调节
胰高血糖素→A激酶→PFK-2失活→F-2,6-P↓↓→抑制氧化\促进糖异生。
胰岛素作用相反。
(二)对PEP―丙酮酸的调节
胰高血糖素→2,6-双磷酸果糖合成↓→1,6-双磷酸果糖合成↓→PK活性↓
胰高血糖素→A激酶→PK磷酸化活性↓↓
胰高血糖素→通过cAMP→诱导PEP羧激酶基因表达(胰岛素与之拮抗)

三、糖异生的意义
(一)维持血糖浓度恒定
1、运动(肌肉收缩)→乳酸→血运输到肝→葡萄糖
2、饥饿→氨基酸(主要)、甘油→葡萄糖
(二)补充肝糖原、恢复糖原储备
(三)调节酸碱平衡
长期饥饿(体液pH降低)→肾PEP羧激酶合成加强→肾糖异生加强→α-酮戊二酸减少→Gln脱氨、排氨入肾小管腔与原尿中H+结合→防止酸中毒。
四、乳酸(Cori)循环
肌肉糖酵解生成的乳酸不能在肌肉组织异生成葡萄糖,必须经血液循环运输至肝脏经糖异生途径生成葡萄糖。原因是肌肉无葡萄糖-6磷酸酶,其它酶活力也低。
乳酸循环的意义:
①避免损失乳酸;②防止因乳酸堆积造成的酸中毒。
乳酸循环是消耗能量的过程:2分子乳酸→1分子葡萄糖;消耗6分子ATP。