
第七章 生物氧化
1、生物氧化(biological oxidation):物质在体内进行氧化称生物氧化。
主要指营养物质在体内分解时逐步释放能量,最终生成CO2和水的过程。
生物氧化又称组织呼吸或细胞呼吸。
生物氧化释放的能量:主要(40%以上)用于ADP的磷酸化生成ATP,供生命活动之需。
其余以热能形式散发用于维持体温。
2、生物氧化内容
(1)生物体内代谢物的氧化作用、代谢物脱下的氢与氧结合成水的过程。
(2)生物体内二氧化碳的生成。
(3)能量的释放、储存、利用(ATP的代谢――ATP的生成与利用)。
3、生物氧化的方式――遵循一般氧化还原规律。
(1)失电子:代谢物的原子或离子在代谢中失去电子,其原子正价升高、负价降低都是氧化。
(2)脱氢:代谢物脱氢原子(H=H++e)的同时失去电子。
(3)加氧:向底物分子直接加入氧原子或氧分子的反应使代谢物价位升高,属于氧化反应。
向底物分子加水、脱氢反应的结果是向底物分子加入氧原子,也属于氧化反应。
4、生物氧化的特点
(1)在温和条件下进行(37℃,中性pH等);
(2)在一系列酶催化下完成;
(3)能量逐步释放,部分储存在ATP分子中;
(4)广泛以加水脱氢方式使物质间接获得氧;
(5)水的生成由脱下的氢与氧结合产生;
(6)反应在有水环境进行;
(7)CO2由有机酸脱羧方式产生。
5、物质体外氧化(燃烧)与生物氧化的比较
(1)物质体内、体外氧化的相同点:
物质在体内外氧化所消耗的氧量、最终产物、和释放的能量均相同。
(2)物质体内、体外氧化的区别:
体外氧化(燃烧)产生的二氧化碳、水由物质中的碳和氢直接与氧结合生成;
能量的释放是瞬间突然释放。
5、营养物氧化的共同规律
糖类、脂类和蛋白质这三大营养物的氧化分解都经历三阶段:
分解成各自的构件分子(组成单位)、降解为乙酰CoA、三羧酸循环。
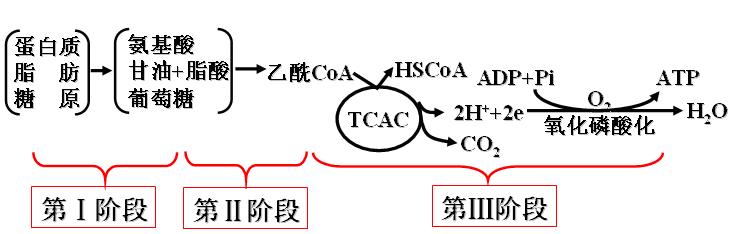
第一节 ATP生成的体系
一、呼吸链( respiratory chain):
代谢物脱下的氢原子(2H)通过多种酶和辅酶所催化的连锁反应逐步传递,最终与氧结合生成水。这一传递链称呼吸链。又称电子传递链(electron transport chain )。
呼吸链由按一定顺序排列在线粒体内膜上的递氢体、递电子体组成。
(一)呼吸链的组成
用胆酸或脱氧胆酸处理线粒体内膜,可将呼吸链分离为:
四种具有递电子功能的酶复合体:
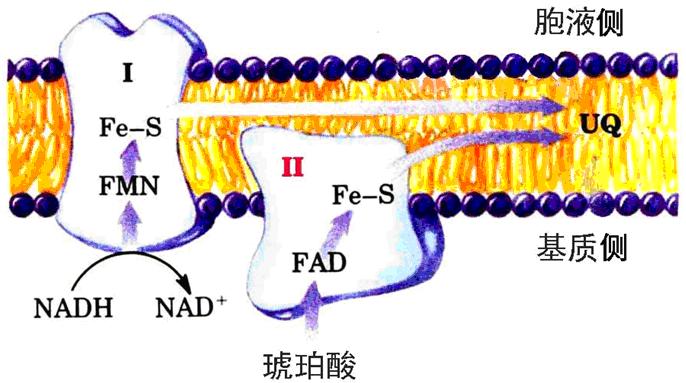
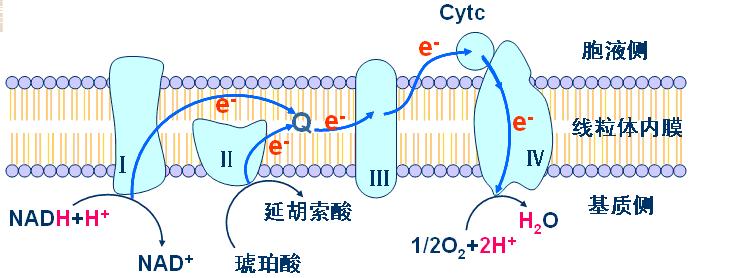
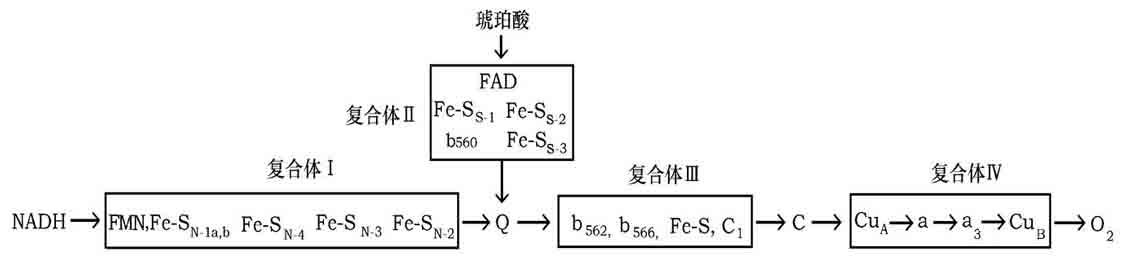
复合体Ⅰ――称NADH一泛醌还原酶(NADH脱氢酶):
复合体Ⅱ――称琥珀酸-泛醌还原酶(琥珀酸脱氢酶):
复合体Ⅲ――称泛醌-细胞色素C还原酶:
复合体Ⅳ――称细胞色素氧化酶(细胞色素aa3):

二种游离成分:
辅酶Q和细胞色素C不包含在复合体中。
1、复合体Ⅰ――将电子从NADH传给CoQ。

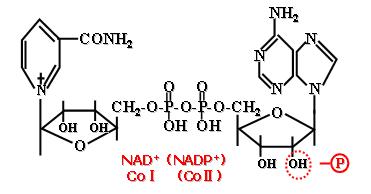
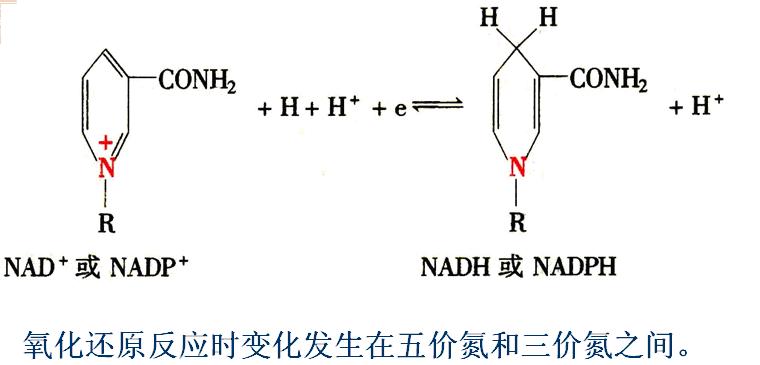
NAD+中的烟酰胺氮为5价,能接受电子成3价,而其对侧碳原子能进行加氢反应(仅1个氢原子)。
(1)NAD+(尼克酰胺腺嘌呤二核苷酸)和NADP+(尼克酰胺腺嘌呤二核苷酸磷酸)
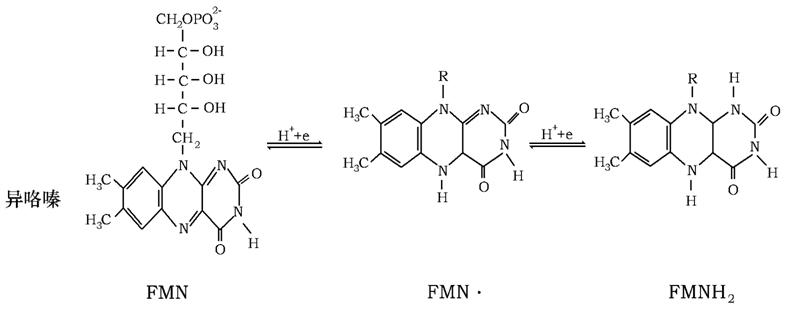
(2)FMN的氧化与还原
黄素单核苷酸(FMN )含有核黄素(维生素B2),FMN中发挥作用的是异咯嗪环。
核黄素与酶的辅基的形成:
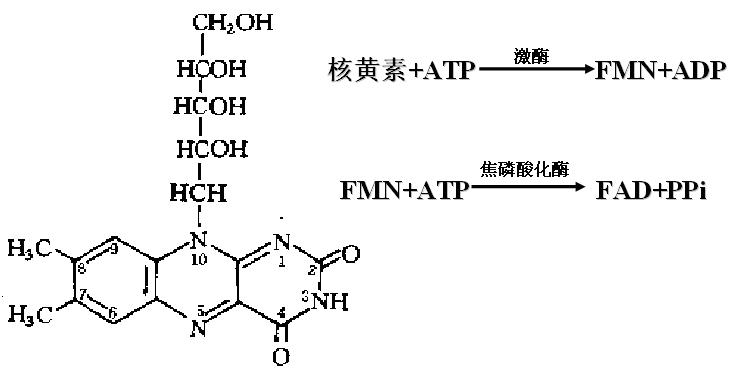
核黄素氧化还原机制:
(3)铁硫蛋白与铁硫中心(Fe-S)
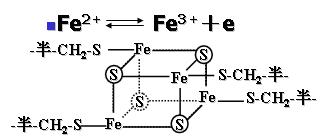
Fe-S含有等量(各4)的铁原子与硫原子;通过铁原子与铁硫蛋白中的半胱氨酸残基的硫相连。
其铁原子可以进行氧化还原反应传递电子。
(4)复合体Ⅰ的功能

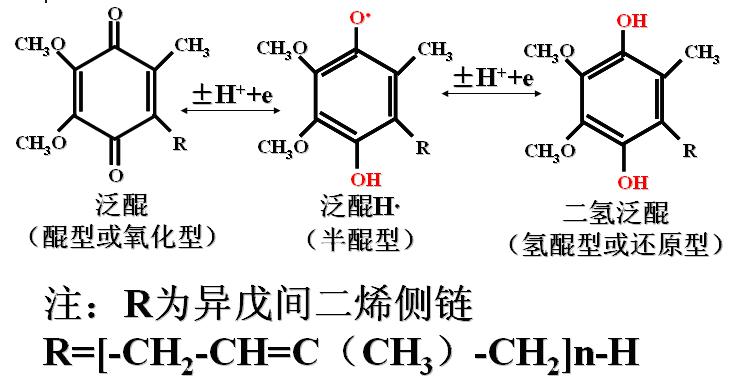
(4)泛醌(CoQ)
辅酶Q是一种脂溶性醌类化合物;含有一个较长的侧链(R),疏水性强。
其侧链由多个异戊间二烯组成,人的CoQ由10个异戊间二烯组成(Q10)。
泛醌可以接受或脱去1个电子和1个质子参与氧化还原。
泛醌参与氧化还原机制:
2、复合体Ⅱ――将电子从琥珀酸传递给CoQ。含如下成分:
含黄素蛋白(以FAD为辅基)、铁硫蛋白(Fe-S)、细胞色素b560。

细胞色素是一类催化电子传递的酶,以铁卟啉为辅基,均有特殊的吸收光谱而呈颜色。
根据吸收光谱的不同,参与呼吸链的Cyt有3类:Cyt a、Cyt b、Cyt c;根据最大吸收峰的差别分为若干亚类。
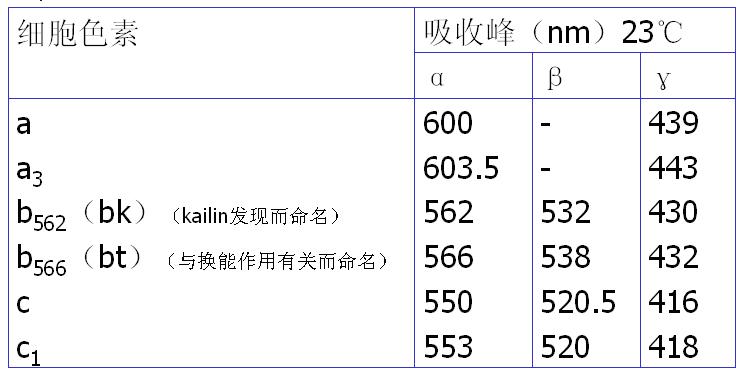
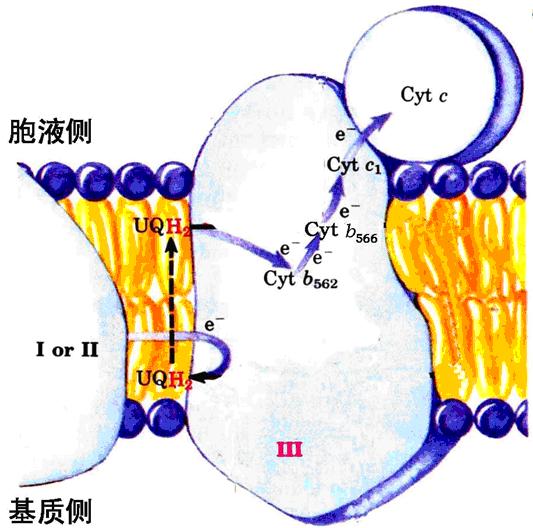
3、复合体Ⅲ――将电子从泛醌传递给细胞色素C。

2种细胞色素b(Cyt b562、 Cyt b566)、细胞色素c1、铁硫蛋白。
细胞色素c呈水溶性,与线粒体内膜外表面结合不紧密而极易于分离,不属于任何复合体成分。
4、复合体Ⅳ――将电子从细胞色素C传递给氧。

含如下成分:细胞色素a、a3 和2个铁卟啉辅基及与之相连的Cu。Cyta和Cyta3很难分开,合称为Cytaa3。其中2个铜原子分别连接2个铁卟啉辅基,铜原子可以进行氧化还原反应传递电子。

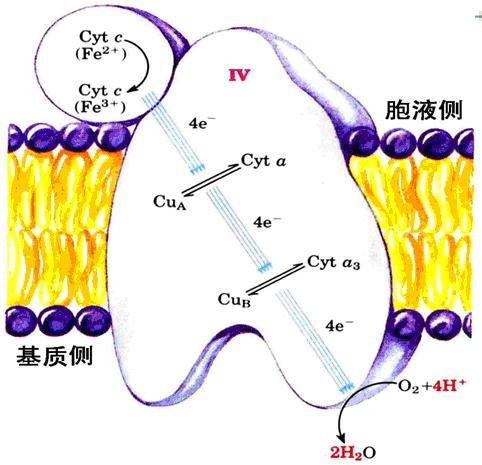
呼吸链各复合体位置示意图
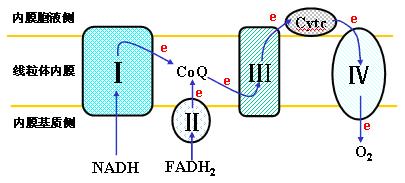
(二)呼吸链成分的排列顺序――确定呼吸链排列顺序的方法:
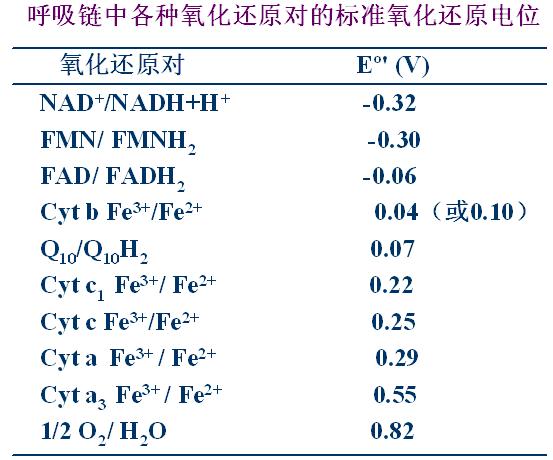
1、根据各组分的EO′从低到高排列――电位低容易失去电子(见下表)。
2、在体外将呼吸链拆开重组――鉴定4种复合体的组成与排列顺序。
3、利用呼吸链特异阻断剂阻断电子传递――阻断部位以前的处于还原状态,后面的组分处于氧化状态,根据吸收光谱的改变检测。
4、以离体线粒体的还原态(无氧)对照――缓慢给氧,通过光谱观察各组分的氧化顺序。(1)氧化-还原对――参加氧化还原反应的每种物质都有氧化型和还原型两种形式:
如:AH2十B←→A十BH2;式中AH2/或BH2是还原型(剂),A和B是氧化型(剂)。
一物质的氧化型/还原型(如A/AH2或B/BH2等)构成氧化还原对(简称氧-还对),氧化还原对是共扼的。氧-还对供出电子趋势的大小,可用标准氧化还原电位Eo′表示。
(2)标准氧化还原电位(Eo′)――成对的氧化型/还原型物质的浓度为1摩尔,在PH7.0,25℃时组成半电池,以(生物化学)规定的标准氢电极做参比测得的电位(伏特/摩尔)。
Eo′值表示的是同H+/H2相比,某氧-还对氧化还原能力的大小:
1)Eo′是负值,表示此氧-还对易供出电子而被氧化,是还原剂;
2)Eo′是正值,表示此氧-还对易获得电子而被还原,是氧化剂。
(3)标准氢电极与生物化学规定的标准氢电极
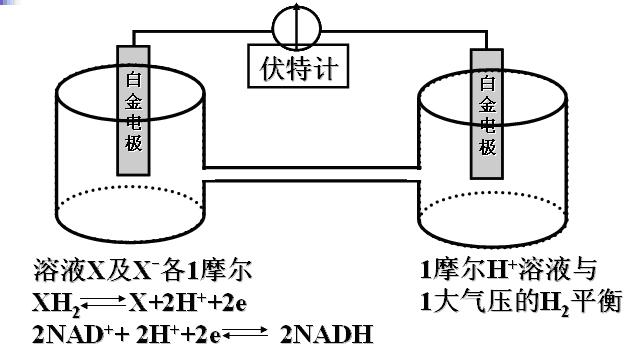
1)标准氢电极:白金电极放入氢离子浓度为1摩尔/升的溶液,与1大气压的氢气平衡,此电极电位定为0伏特,作为参比电极Eo。
2)生物化学上规定:加入pH=7.0的条件,将氢电极浓度定为10-7mol/L,测得的电位是Eo′。
(4)电位测定装置
2、两条氧化呼吸链及其排列顺序
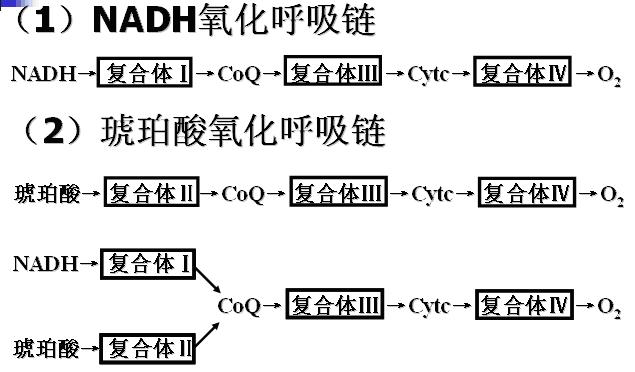
3、呼吸链电子传递过程

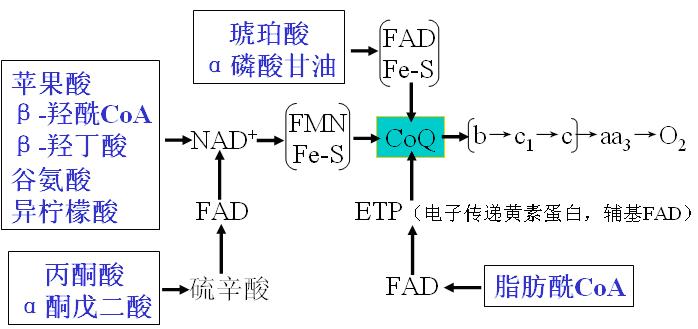
4、重要代谢物进入呼吸链的途径
二、氧化磷酸化
ATP是机体主要供能物质。
ATP形成的主要方式是氧化磷酸化:呼吸链电子传递过程中偶联ADP磷酸化生成ATP。
ATP的另外形成方式是底物水平磷酸化:直接将代谢物分子中的能量转移到ADP(GDP)生成ATP(GTP)。
(一)氧化磷酸化的偶联部位――ATP生成的部位
偶联部位的确定方法如下:
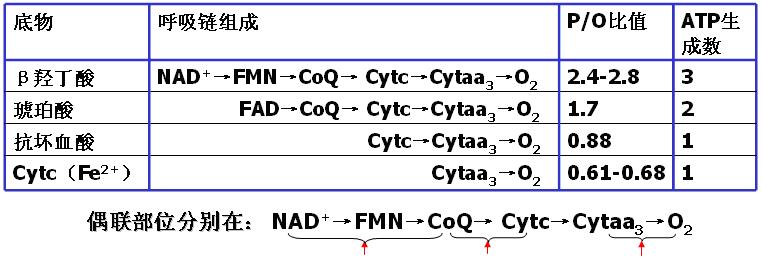
1、P/O比值:物质氧化过程中每消耗1摩尔氧原子所消耗的无机磷或ADP的摩尔数。
此数值代表ATP的生成数。P/O的测定方法:
将底物、ADP、H3PO4、Mg2+和分离较为完整的线粒体在模拟细胞内液的环境中相互作用;测定O2和H3PO4(或ADP)的消耗量,可以计算P/O,确定偶联部位。
2、自由能变化
ΔGo′=- nFΔEo′>30.5kJ可合成ATP,ΔGo′表示pH7时的标准自由能变化, n=2个氧还对反应时传递电子的数目(2),F是法拉第常数(96.5kJ/mol?V),ΔEo′为电位变化。
自由能变化的计算举例――NADH(H+)的氧化反应:
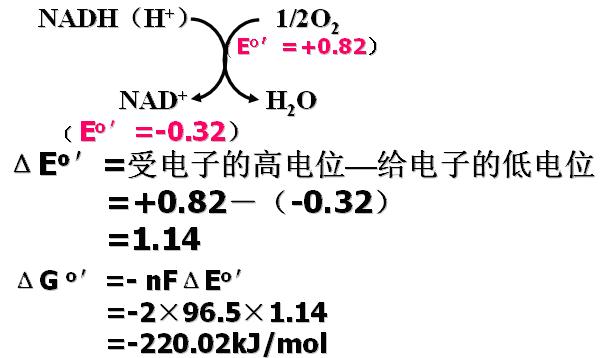
从电位差( ΔEo′)计算ΔGo′,如果ΔGo′足以达到合成ATP(释放的自由能>30.5kJ/mol)则存在偶联部位。30.5kJ是合成1molATP所需的能量(标准状态),体内条件ΔGo′=-51.6kJ/mol
(二)氧化磷酸化偶联机制――化学渗透学说
化学渗透学说要点:
1、呼吸链中递氢体与递电子体交替排列,并在膜中有固定位置,催化的反应是定向的,取决于电子走向。
2、电子经呼吸链传递时可将质子从线粒体内膜的基质泵到内膜外侧,产生膜内外质子电化学梯度(氢离子浓度梯度和跨膜电位差),以此储存能量。
3、当质子顺浓度梯度回流时驱动ATP合酶,利用ADP和Pi合成ATP。
说明:
电子传递链在线粒体内膜中共构成3个回路,即形成3个氧化还原袢,每个回路均有质子泵。
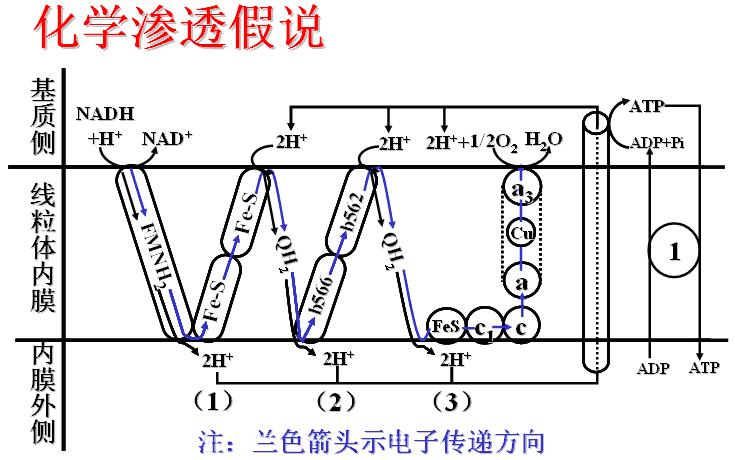
呼吸链模式
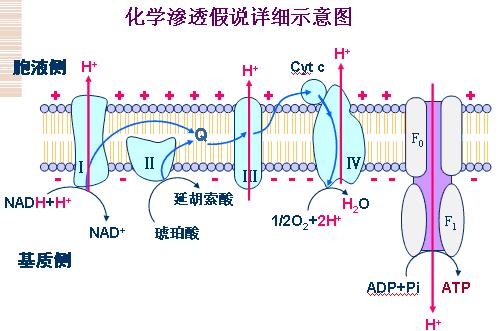
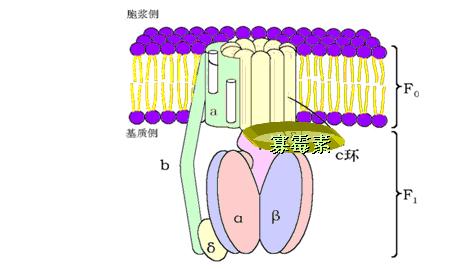
(三)ATP合酶(复合体Ⅴ)
ATP合酶(ATP synthase)――三联体:
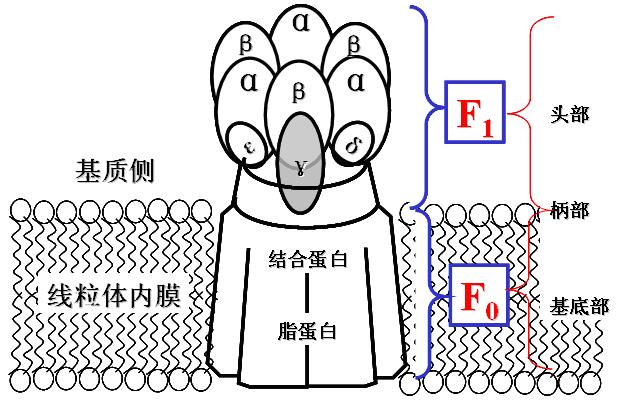
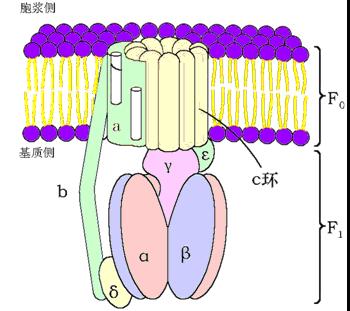
F1:亲水部分,位于线粒体内膜的基质侧,含有5种肽链、9个甲基(α3β3γδε)。
功能是催化ATP的生成。催化部位在β亚基,但必须与α亚基结合才有活性。
F0:疏水部分,镶嵌在线粒体内膜中的H+通道。
当H+顺浓度梯度经F0回流时,F1催化ADP和Pi合成并释放ATP。
F0和F1之间有寡霉素敏感蛋白(OSCP),OSCP使ATP合酶在寡霉素存在时无作用。
线粒体结构
1、ATP合酶的结构模式
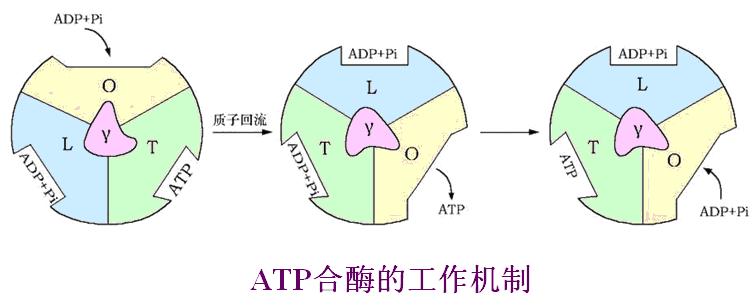
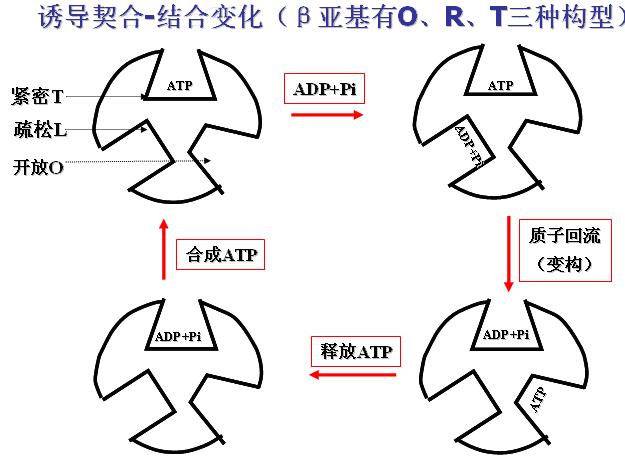
2、ATP的作用机制――诱导契合-结合变化(β亚基有O、R、T三种构型)
三、影响氧化磷酸化的因素
氧化磷酸化的影响因素有:
呼吸链抑制剂、解偶联剂、氧化磷酸化抑制剂;ADP的调节作用、甲状腺素的作用、线粒体DNA突变。
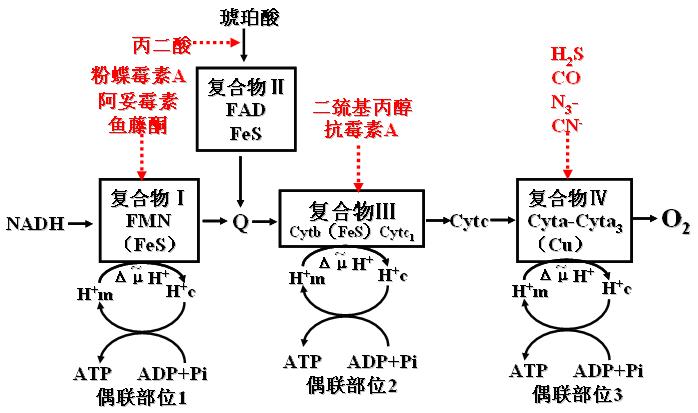
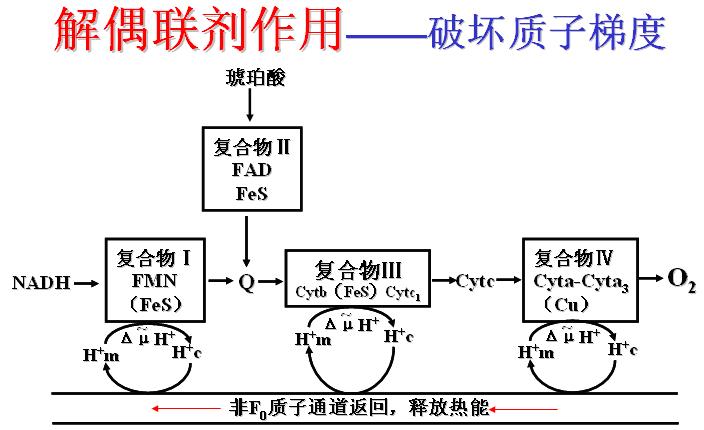
(一)呼吸链抑制剂,阻断电子传递。此抑制剂可以停止细胞内呼吸,引起死亡。
(1)鱼藤酮、粉蝶霉素A、异戊巴比妥――与复合体Ⅰ的铁硫蛋白结合而阻断电子传递。
(2)抗霉素A、二巯基丙醇(BAL)――抑制复合体Ⅲ中Cytb与Cytc1间的电子传递。
(3)CO、CN-、N3-、H2S――抑制复合体Ⅳ(细胞色素氧化酶),使电子不能传递给氧。
呼吸链抑制剂及部位
(二)解偶联剂(多属于能通过膜的阳离子 载体),使氧化与磷酸化过程脱离。
作用机制:使呼吸链电子传递过程泵出的氢离子不经过ATP合酶的F0质子通道回流,而经其他途径返回线粒体基质。破坏膜两侧的电化学梯度,电化学梯度储存的能量以热能形式散发。
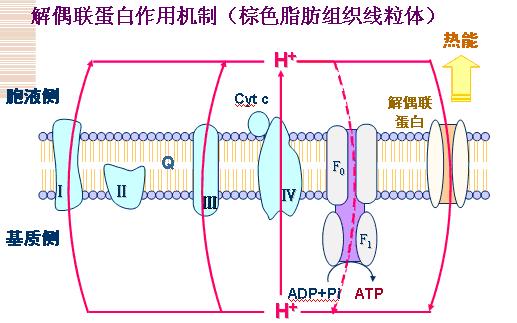
1、 解偶联蛋白质子通道(释放热能)存在于动物棕色脂肪组织;
2、 FFA促进质子经解偶联蛋白反流至基质。
3、 二硝基苯酚(DNP)结合质子在膜内移动。
4、 其他:游离脂肪酸、水杨酸盐、双香豆素。
(三)氧化磷酸化抑制剂――对电子传递与ADP的磷酸化均抑制。
如寡霉素可与ATP合酶F1、F0之间柄部的寡霉素敏感蛋白(OSCP)结合,阻止质子从F0通道内流合成ATP。质子不能内流导致膜两侧电化学梯度增高,影响质子泵的功能,进而抑制电子传递。
如苍术苷(酸):特异抑制ATP/ADP载体(腺苷酸转位酶)。
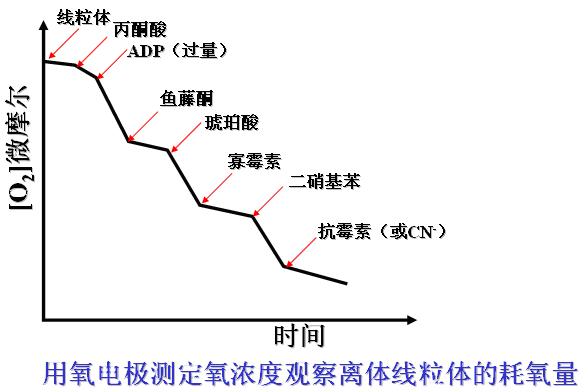
各种抑制剂对线粒体耗氧量的影响――实验过程:
(二)ADP的调节作用
正常机体氧化磷酸化的速率主要受ADP的调节:
机体利用ATP ↑ →ADP ↑ →ADP进入线粒体↑ →氧化磷酸化↑。
反之ADP不足→氧化磷酸化↑。
这种调节可使ATP的生成适应生理需要。
用极谱法氧电极系统测量游离线粒体的呼吸过程
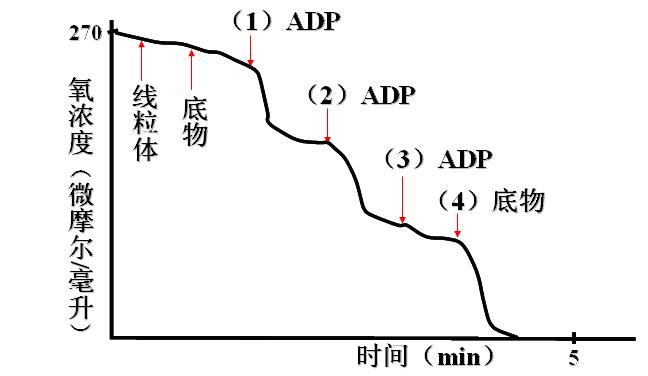
(三)甲状腺素
甲状腺素的作用有两个方面:
1、促进氧化磷酸化:甲状腺素诱导膜上Na+-K+-ATP酶的合成,促进ATP分解为ADP,而促进氧化磷酸化。
2、甲状腺素(T3)可以增加解偶联蛋白的表达,引起耗氧合产热均增加,即基础代谢率增加。
(四)线粒体DNA突变
mtDNA是裸露的环状双螺旋,缺乏蛋白质的保护和损伤修复系统,容易受本身氧化磷酸化过程产生的氧自由基的损伤而发生突变。突变率是核内DNA的10~20倍。
mtDNA编码的基因:呼吸链中13条肽链、线粒体蛋白合成所需要的22个tRNA、蛋白合成所需要的2个rRNA。
mtDNA突变可以降低线粒体的功能。mtDNA病的主要问题是ATP 生成减少引起的症状,耗能较多的器官先受累,且随年龄的增长而严重。
四、ATP――能量的储存的形式
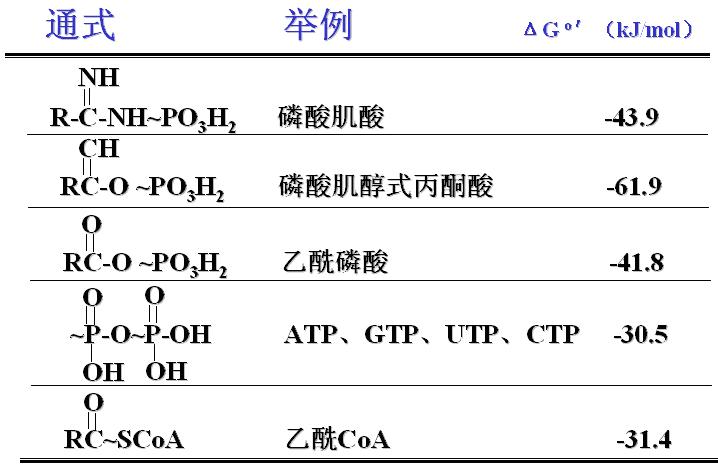
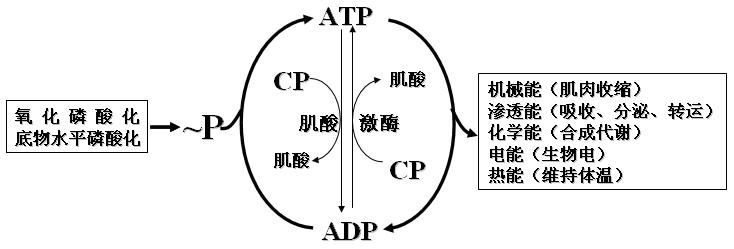
生物氧化过程释放的能量有约40%以化学能的形式储存于特殊的有机磷酸化合物中,形成磷酸酯(磷酸酐)。磷酸酯键水解放能较多大于21kJ/mol,称为“高能磷酸键”,用“~P”表示。
在所有高能磷酸化合物中,以ATP分子末端的γ磷酸键最为重要。
(一)常见的高能磷酸化合物
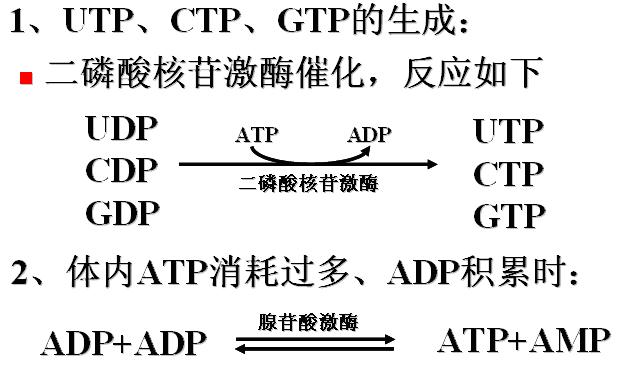
(二)各种核苷酸之间的转变
(三)磷酸肌酸
1、ATP可以将~P转移给肌酸生成磷酸肌酸。反应可逆。过程如下:
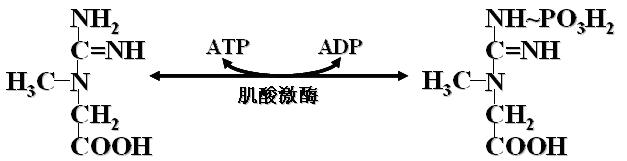
2、磷酸肌酸是肌肉和脑组织能量的一种储存形式。
(四)ATP的生成和利用――生物体能量的生成和利用都是以ATP为中心。
体外pH7.0、25℃条件下,1摩尔ATP水解为ADP+Pi时释放的能量为30.5kJ,体内生理条件下为50kJ/mol。人体每日经ADP/ATP相互转变的量(ATP转换率)非常可观。
静卧24小时消耗ATP40kg,运动时0.5kg/min。
1、ATP的利用――ATP循环(细胞能量循环)
(1)末端磷酸基的分裂与转移,生成ADP和新的磷酸化物。
如激酶催化的反应。此时ATP的磷酸基和部分能量同时转给新的磷酸化物。
(2)ATP水解为ADP+Pi,能量供机体利用。
离子转运、肌肉收缩、羧化反应。
(3)利用ATP的另一个高能磷酸键。
生成焦磷酸。如脂酸活化反应。
2、ATP生成、储存和利用
五、通过线粒体内膜的物质转运
线粒体基质与细胞液之间有线粒体内外膜相隔。
线粒体外膜通透性好,允许分子量在1000以内的物质自由通过(普通生物膜)。
线粒体内膜对各种物质的通过有严格选择性。
几乎所有的离子、不带电荷的小分子有机化合物都不能自由通过,必须依赖内膜上特殊的蛋白。
此蛋白少数是酶,多数是载体(也称为转位酶)。载体对物质具有严格的选择性。
内膜载体对物质的转运规律:一种离子或带电荷的化合物顺浓度梯度向内膜一侧转运时,必须伴有对应离子或带电物质逆浓度梯度反向转运。通常是“一对一交换”(A-入B-出或A-出B-入)。
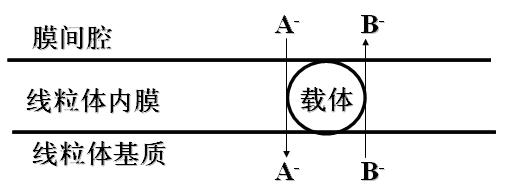
线粒体内膜的运载体(重要的线粒体转位酶)
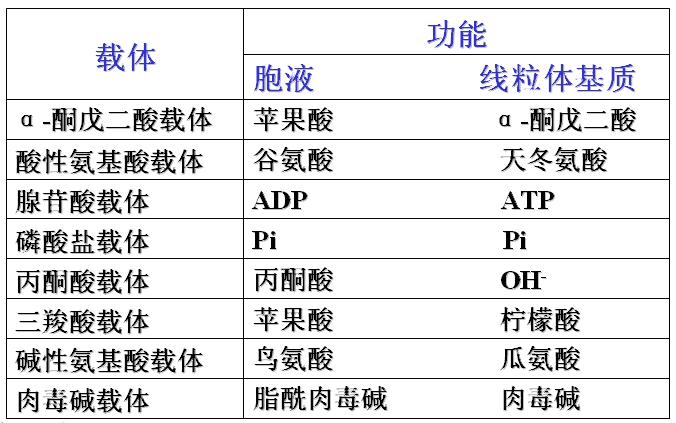
四种转位酶的协同作用
(一)胞液中NADH的氧化――如何进入线粒体
胞液脱氢产生的还原当量(H++e)必须进入线粒体氧化,但NADH不能自由透过线粒体内膜。必须借助于其他转运机制完成。有两种机制可以使NADH进入线粒体:α-磷酸甘油穿梭和苹果酸-天冬氨酸穿梭。
1、α-磷酸甘油穿梭――存在于脑和骨骼肌
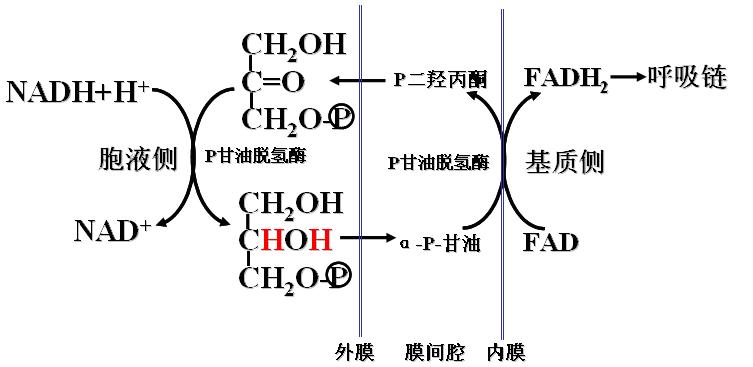
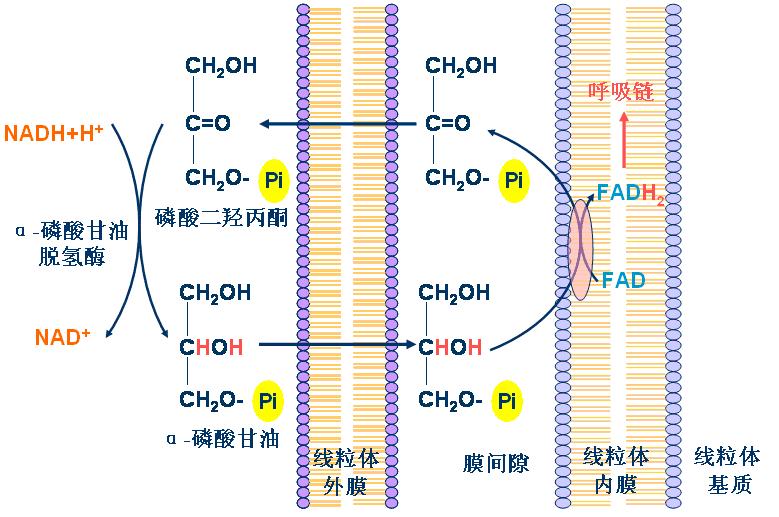
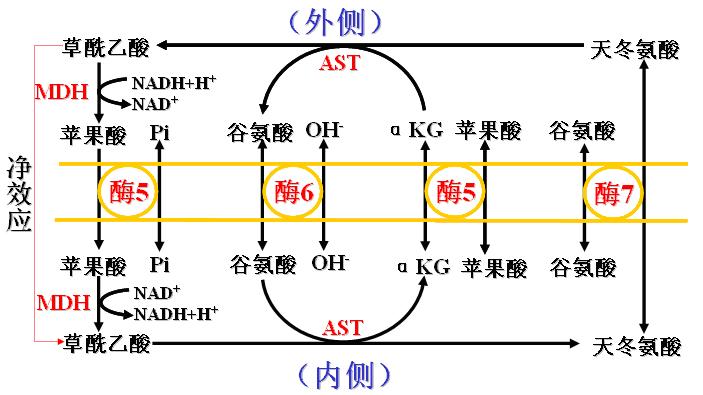
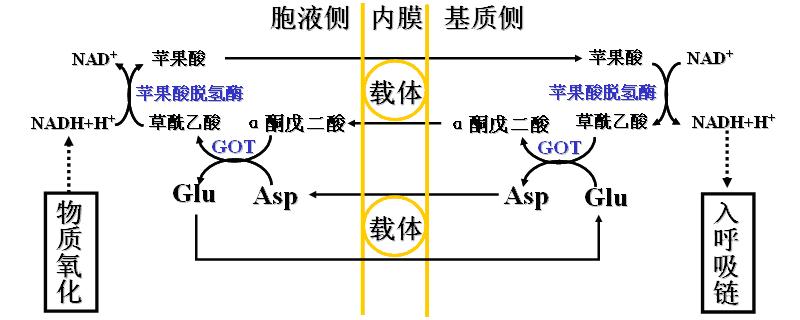
2、苹果酸-天冬氨酸穿梭――存在于肝脏和心肌
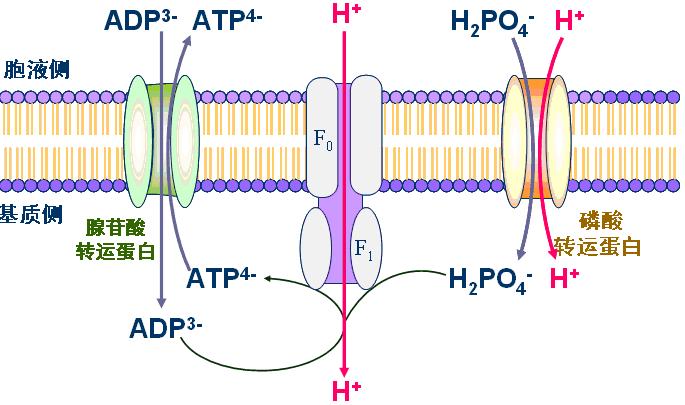
(二)腺苷酸载体――又称ATP-ADP载体或ATP-ADP转位酶。
2个亚基组成的二聚体,每个亚基3.0kD。ATP与ADP经此载体反向交换;同时胞液中的H2PO4-经磷酸盐载体与H+同向转运入线粒体。
ATP、ADP、Pi的转运

(三)线粒体蛋白质的跨膜转运
线粒体蛋白90%是由核DNA编码的,在线粒体外合成。
1、基质内蛋白质:
蛋白质前体肽链在外膜表面由解折叠酶使其空间结构松散;并被外膜上的受体识别后转移到总插入蛋白;总插入蛋白使蛋白前体从氨基端开始进入基质;基质中的加工肽酶切除蛋白前体中的导向信息肽段;形成成熟的基质蛋白质。
2、线粒体内膜或膜间隙的蛋白质:
基质中加工肽酶作用后暴露新的氨基端疏水肽段;疏水肽段引导肽连重新穿过内膜;间隙中的酶切除疏水肽段生成成熟蛋白质。
第二节 其他氧化体系
一、需氧脱氢酶和氧化酶:直接以氧为受氢体。
(一)需氧脱氢酶(辅基是FMN或FAD),产物是过氧化氢。
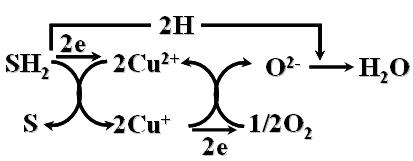
(二)氧化酶(辅基含有铜离子),产物中有水。氧化过程如下:
二、过氧化物酶体中的氧化酶
(一)过氧化氢酶(触酶):辅基含4个血红素
反应如下: 2H2O2 → 2H2O + O2
在粒细胞和吞噬细胞中H2O2可以杀死入侵的细菌;
甲状腺细胞中H2O2可使2I-氧化为I2,使酪氨酸碘化成甲状腺素(T3、4)。
(二)过氧化物酶:也含有血红素辅基。
催化H2O2直接氧化酚类和胺类。反应如下:
R + H2O2 → RO + H2O 或者 RH2 + H2O2 →R + 2H2O
三、超氧化物歧化酶(SOD)
呼吸链电子传递过程可以产生超氧离子;其他物质(如黄嘌呤)氧化也可以产生超氧离子。
超氧离子可进一步生成H2O2和羟自由基(?OH),统称为反应氧族。
反应氧族化学性质活泼,可以使磷脂中的不饱和脂肪酸氧化成过氧化脂质,损伤生物膜;过氧化脂质与蛋白质结合成的复合物积累成的棕褐色色素颗粒即脂褐素,与组织老化有关。
超氧化物歧化酶(SOD)可以消除超氧离子:
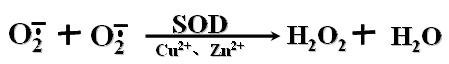
四、微粒体中的氧化酶
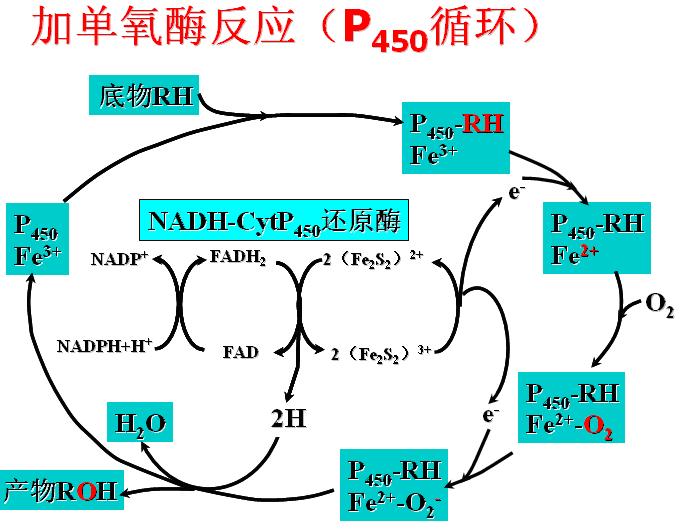
(一)加单氧酶(羟化酶、混合功能氧化酶)
反应如下:
R+NADPH+H++O2 → ROH+NADP++ H2O
反应需要细胞色素P450(与CO结合后在450nm有最大吸收峰,故名)参加。
(二)加双氧酶
催化O2的2个氧原子加到带双键的两个碳原子上。
